


{kind=link}
{kind=link}
{kind=link}
宁夏荒漠草原不同群落蒙古冰草种群空间格局及种间关联性
[许爱云1  , 许冬梅
, 许冬梅1, 2, * , 曹兵1 , 刘金龙1 , 于双1 , 郭艳菊1 , 马晓静1 ]
, 许冬梅, 曹兵|
|


作者简介:许爱云(1994-),女,甘肃庆阳人,在读博士。E-mail: xaynxu@163.com
为探讨蒙古冰草种群对环境的适应对策,以宁夏盐池县荒漠草原短花针茅+蒙古冰草群落、蒙古冰草+牛枝子群落、蒙古冰草+牛心朴子群落为研究对象,采用点格局分析中的O-ring函数统计方法,分析了不同群落中蒙古冰草种群的数量特征、分布类型及优势种群的种间关联性。结果表明:不同群落生境条件下,蒙古冰草+牛枝子群落中蒙古冰草种群的密度、盖度和地上生物量显著高于短花针茅+蒙古冰草群落和蒙古冰草+牛心朴子群落( P<0.05),3个群落之间蒙古冰草种群的高度差异不显著( P<0.05)。不同群落生境蒙古冰草种群在<4 m尺度范围内主要表现为聚集分布,随尺度增大,聚集强度减弱,逐渐过渡到随机分布和均匀分布。蒙古冰草种群与短花针茅种群、蒙古冰草种群与牛枝子种群在小于4 m尺度上呈显著负关联,随尺度的增大趋于无关联;蒙古冰草种群与牛心朴子种群在整个研究尺度上无关联。
Population characteristics and interspecies relationships are important in the study of population ecology. To investigate the environmental plasticity mechanisms of Agropyron mongolicum populations, three 10 m×10 m plots dominated by communities of Stipa breviflora+ A. mongolicum, A. mongolicum+ Lespedeza potaninii and A. mongolicum+ Cynanchum komarovii were studied in desert steppe grassland in Yanchi County. The spatial coordinates and quantitative characteristics, including height and above ground biomass of A. mongolicum were measured in the study plots. The spatial distribution pattern of the A. mongolicum populations and the interspecific relationships between the A. mongolicum populations and other dominant species in the studied plant communities were analyzed by ‘point pattern analysis’ and ‘O-ring function statistic’ methods. It was found that the density, ground cover and above-ground biomass of A. mongolicum populations in A. mongolicum+ L. potaninii communities were significantly higher than those in S. breviflora+ A. mongolicum and A. mongolicum+ C. komarovii communities ( P<0.05), and the height did not differ significantly among the three communities ( P<0.05). The spatial pattern of A. mongolicum populations mainly displayed an aggregated distribution, with an aggregation scale of less than 4 m. When spatial distribution was evaluated at larger scale, the aggregation degree decreased with increasing scale and gradually transformed to random and uniform distribution. The associations between A. mongolicum and S. breviflora populations, and A. mongolicum and L. potaninii populations showed a negative relationship at a scale of less than 4 m and tended to show no association with increase in the evaluation scale. No association between A. mongolicum and C. komarovii populations was detected at the 0-5 m evaluation scale.
种群是在一定空间和时间内同种生物个体的集合[1]。种群空间格局是特定时间内群落中种群个体在水平空间上的配置特点, 一定程度上反映了物种与生境的关系及其在群落中的地位与作用[2]。为适应水分、养分与光照等生境条件, 植物往往会调节自身的形态结构、功能性状与分布策略[3, 4]。植物的高度直接影响光照来源、热量负荷、湿度和种子扩散[5], 较高植物个体可充分利用上层优势获得更多有效资源与空间, 从而抑制相邻个体的生长与分布。干旱条件下, 植物可通过生长高度的改变来调节根际水分到达叶部的距离[4], 同时, 逆境胁迫下, 无性繁殖植物也会通过根茎分蘖产生大量无性分株增加母体适合度[6], 排除具有相同生态位的竞争物种, 最大限度扩展种群领地、提升种群地位[7]。群体效应也是植物适应严酷生境的一种策略[8, 9], 在沙质生境中, 较小个体为了防止被风沙掩埋, 它们彼此合作, 相互庇护, 实现协同生长, 常呈聚集分布[8]; 但随尺度增大, 为争夺有限的群落资源, 种内种间竞争加剧, 植物种群往往由聚集分布逐渐过渡到随机分布和均匀分布, 种间关联性也逐步转为无关联[8, 10, 11]。研究植物种群在不同生境条件下的空间格局, 有助于深入认识种群的生态适应对策、资源利用状况及群落多样性维持机制。
近年来, 随着全球气候的变化, 加之人类的过度干扰, 导致荒漠草原退化, 生态系统稳定性降低[12, 13]。目前, 关于荒漠草原植被和地境方面已开展了大量研究, 主要包括不同恢复措施下土壤理化性状的时空变异[12, 14], 土壤种子库特征[15], 植物群落结构、多样性及稳定性的动态变化[13, 16]等。蒙古冰草(Agropyron mongolicum)又称沙芦草, 为多年生疏丛禾草, 具有较强的抗旱、抗寒、耐风沙等特性, 在我国北方草原区广泛分布, 是宁夏荒漠草原的优势种群之一[15], 作为当地优质牧草, 有其独特的生态过程, 在植被生态修复中具有重要作用[17]。以往对荒漠草原蒙古冰草种群的研究主要集中在土壤种子库[15]、生殖分配[17]及生理特征[18]等方面, 而关于蒙古冰草种群在不同群落的分布类型及与其他优势种群种间关系的研究鲜有报道。本研究选取宁夏荒漠草原短花针茅(Stipa breviflora)+蒙古冰草群落、蒙古冰草+牛枝子(Lespedeza potaninii)群落和蒙古冰草+牛心朴子(Cynanchum komarovii)群落为对象, 研究不同群落蒙古冰草种群的数量特征与分布规律, 以期深入认识蒙古冰草种群生态过程、种间关系及其对环境的适应机制, 为制定有效的荒漠草原生态系统保护和恢复措施提供理论依据。
研究区位于宁夏东部盐池县花马池镇宁夏大学四墩子教学科研基地(107° 15'40″ E, 37° 20'12″ N), 地处毛乌素沙地南缘, 是黄土高原向鄂尔多斯台地的过渡地带, 也是陕、甘、宁、内蒙古四省(区)的交界区域。自然条件较为恶劣, 干旱少雨, 风沙大, 属典型的中温带大陆性季风气候, 年平均气温7.7 ℃, 1和7月平均气温分别为-8.9和22.5 ℃, 年温差较大, ≥ 0 ℃年积温为3430.3 ℃; 年平均降水量289.4 mm, 主要集中在6-8月, 年蒸发量2135 mm, 年无霜期165 d。土壤类型主要为灰钙土、风沙土, 土壤质地为沙壤和粉沙壤, 土壤结构松散, 肥力低下[15]。植被类型属荒漠草原, 主要物种有蒙古冰草、牛枝子、短花针茅、牛心朴子、黑沙蒿(Artemisia ordosica)、赖草(Leymus secalinus)、甘草(Glycyrrhiza uralensis)和猪毛蒿(Artemisia scoparia)等。
于2018年7月中下旬植物生长旺盛时期, 在宁夏大学四墩子教学科研基地分别选取以蒙古冰草种群为优势种或亚优势种的短花针茅+蒙古冰草群落、蒙古冰草+牛枝子群落和蒙古冰草+牛心朴子群落, 在每个群落的中心地段设置1个10 m× 10 m的典型样地。采用邻接格子法, 用1 m× 1 m的样方框按从东到西, 从南到北的顺序依次放置100次进行取样, 以样地的一角顶点为基点(0, 0), 确定每株蒙古冰草及短花针茅、牛枝子和牛心朴子在样地中的二维坐标[19, 20]; 同时, 在每个群落内沿对角线设置9个1 m× 1 m 的样方, 测定蒙古冰草种群的高度、密度、盖度和地上生物量[16]。
按照取样时样方框放置的顺序将100个1 m× 1 m的样方进行拼接, 合并成10 m× 10 m的样地图, 用于空间分布格局与种间关联性分析[19]。
基于蒙古冰草种群空间坐标点信息, 运用点格局分析中的单变量O-ring函数进行蒙古冰草种群不同生境的空间分布格局分析, 双变量O-ring函数分析不同生境种群间的关联性[9, 21]。单变量统计中O-ring函数的O(r)估计值是通过设定对象1等于对象2来计算[9, 21]。双变量O-ring函数的公式为:
Point2[
Area[
式中:n1为双变量统计中对象1的数目;
采用Programita(2014版)软件进行空间二维坐标的分析, Origin 8.0进行绘图, 根据样方大小, 采用的空间尺度为0~5 m, 分析步长0.1 m; 利用Monte-Carlo随机模拟99次, 拟合检验上、下包迹线, 即置信区间为99%。其他数据处理采用Excel 2013与 SPSS 21.0软件完成。
不同群落生境中, 蒙古冰草种群密度、盖度和地上生物量变化显著(表1)。蒙古冰草+牛枝子群落中蒙古冰草种群密度、盖度和地上生物量均显著高于其他两个群落(P< 0.05), 短花针茅+蒙古冰草群落中蒙古冰草种群密度、盖度和地上生物量均最小, 但与蒙古冰草+牛心朴子群落差异不显著(P< 0.05); 蒙古冰草种群高度在3个群落之间无显著差异(P< 0.05)。不同群落优势种群个体的空间分布如图1所示, 可以看出, 蒙古冰草种群个体数量以蒙古冰草+牛枝子群落最多, 蒙古冰草+牛心朴子群落次之, 短花针茅+蒙古冰草群落中蒙古冰草个体数量最少。
| 表1 不同群落蒙古冰草种群数量特征 Table 1 Quantitative characteristics of A.mongolicum population in different plant communities |
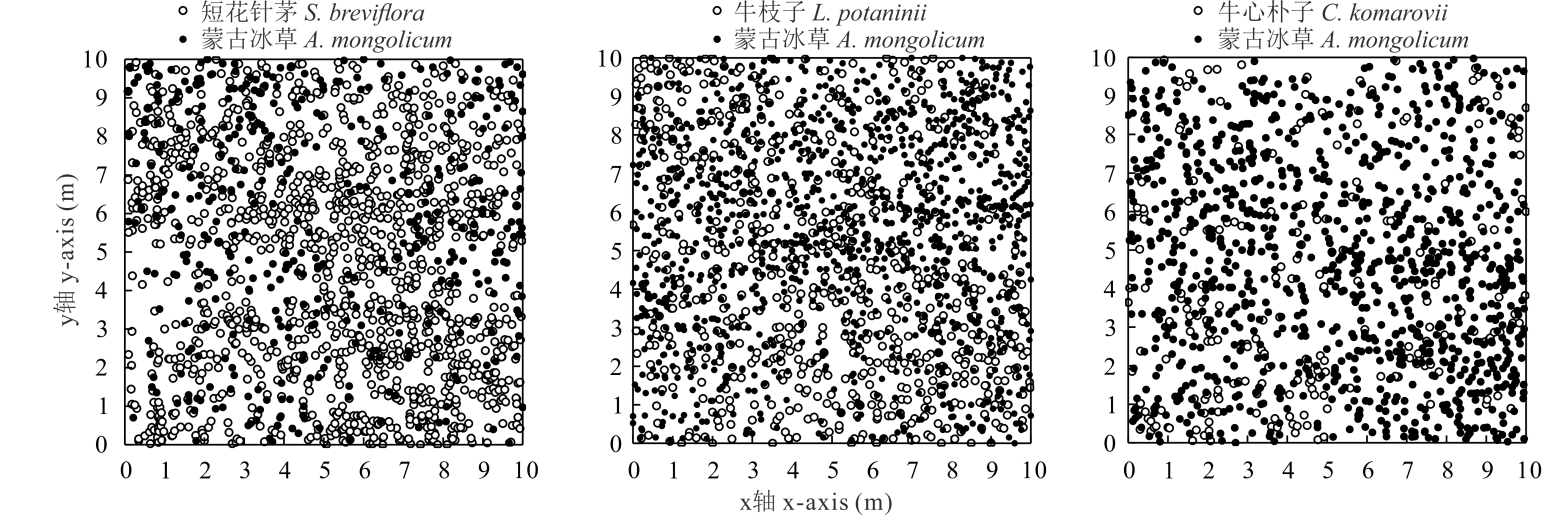 | 图1 不同群落样方中蒙古冰草种群的空间分布Fig.1 Spatial distribution for A. mongolicum population in the sample among different plant communities |
从图2可以看出, 短花针茅+蒙古冰草群落中, 蒙古冰草种群在0.1~1.5 m尺度范围内表现为聚集分布, 在0~0.1 m与1.5~4.6 m范围内表现为随机分布, 在> 4.6 m尺度范围表现为均匀分布; 蒙古冰草+牛枝子群落中, 蒙古冰草种群在0~4.0 m尺度范围内表现为聚集分布, 在> 4.0 m尺度上表现为随机分布; 蒙古冰草+牛心朴子群落中, 蒙古冰草种群在0.2~2.9 m尺度范围内表现为聚集分布, 而在0~0.2 m和2.9~5.0 m尺度范围内表现为随机分布。总体来看, 在< 4.0 m小尺度范围内, 蒙古冰草种群在不同生境群落主要表现为聚集分布, 并且聚集程度较高, 但随着尺度的增大, 聚集强度均明显减弱, 逐渐过渡到随机分布和均匀分布。
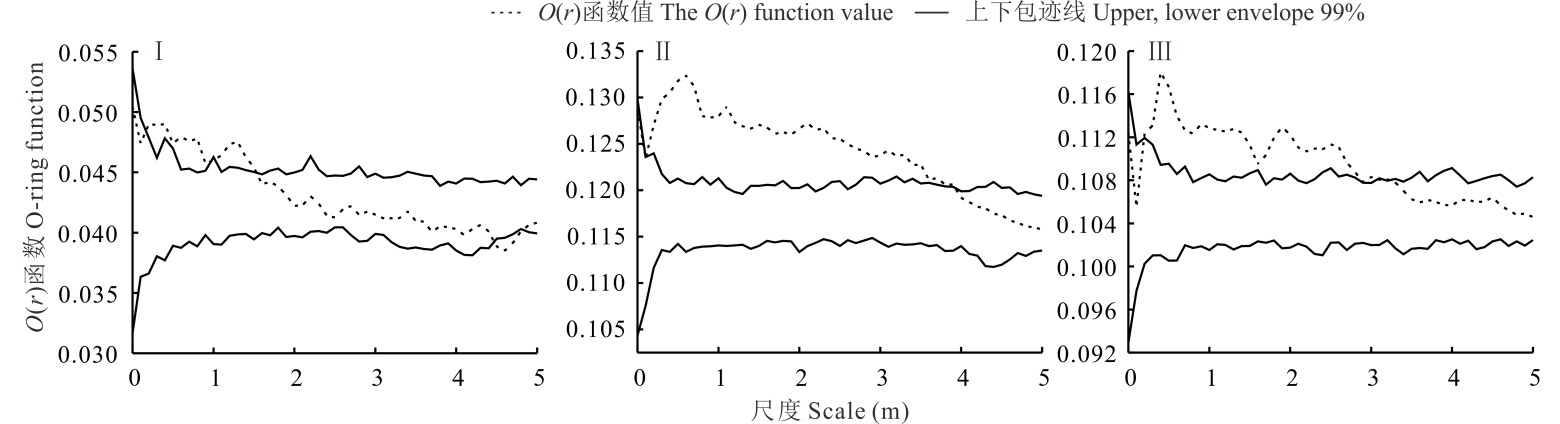 | 图2 不同群落蒙古冰草种群的空间分布格局 Ⅰ 、Ⅱ 、Ⅲ 分别表示短花针茅+蒙古冰草群落、蒙古冰草+牛枝子群落与蒙古冰草+牛心朴子群落中蒙古冰草, 下同。Fig.2 Spatial distribution patterns of A. mongolicum population in different communities Ⅰ , Ⅱ and Ⅲ represent spatial distribution patterns of A. mongolicum population in S. breviflora+A. mongolicum, A. mongolicum+L. potaninii and A. mongolicum+C. komarovii communities respectively, the same below. |
如图3所示, 不同群落蒙古冰草种群与其他优势种群的空间关联性随尺度的变化表现为不同的关联类型。蒙古冰草种群与短花针茅种群在0~1.0 m尺度范围内呈负关联, 在> 1.0 m尺度上无关联; 蒙古冰草种群与牛枝子种群在0~2.9 m尺度上呈负关联, 在> 2.9 m尺度上无关联; 而蒙古冰草种群与牛心朴子种群之间在整个研究尺度范围内均表现为无关联性。
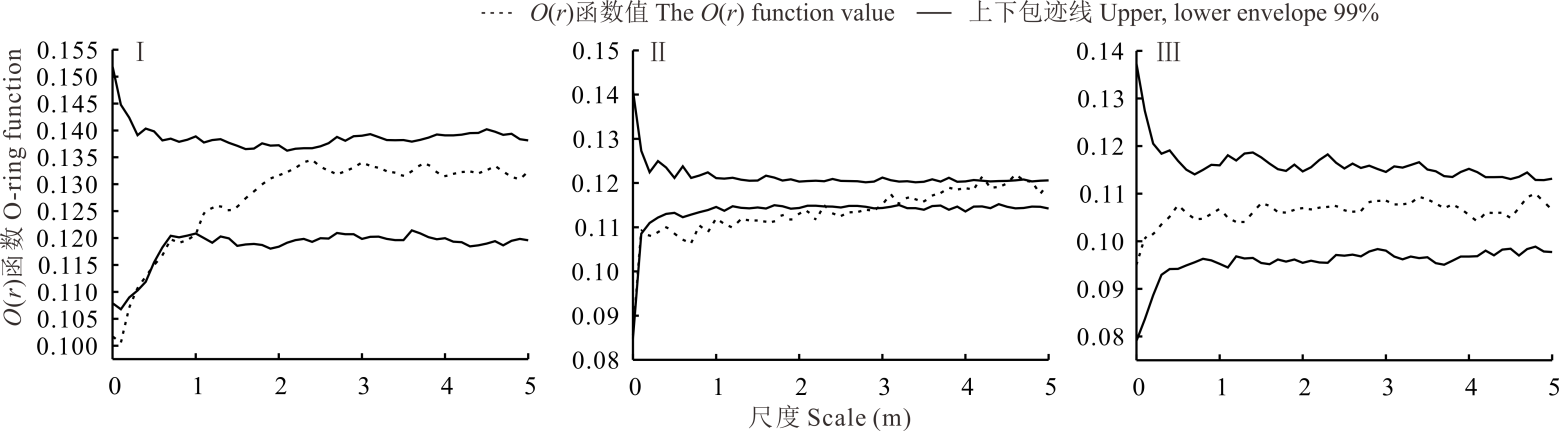 | 图3 不同群落蒙古冰草与其他种群的空间关联性Fig.3 Spatial association between A. mongolicum and other population in different communities |
植物数量特征与分布规律是植物种内种间关系对群落生境的响应, 可反映种群在群落中的地位及资源利用状况[7, 22]。3个群落之间蒙古冰草种群的密度、盖度和地上生物量差异显著, 表明蒙古冰草一方面通过个体性状的可塑性变化优化资源配置, 另一方面通过种群数量特征的改变调节种内种间关系, 以适应不同的生境条件[22]。随尺度增大, 各群落生境蒙古冰草种群由聚集分布逐渐过渡到随机分布和均匀分布。短花针茅+蒙古冰草群落中蒙古冰草种群仅在0.1~1.5 m尺度范围内呈聚集分布, 且聚集强度小于其他两个群落。出现该结果的可能原因有:1) 植物的生物学特性。聚集分布是植物适应生境变化的一种表现[8, 9]。种群在小尺度内呈聚集分布, 有利于其发挥群体效应, 提高种群竞争力, 维持种群的正常发展[8]。研究区生境条件严酷[12, 16], 为了提高存活机会, 蒙古冰草种群内个体间彼此合作, 相互庇护, 进而表现出聚集分布的特征。随尺度增大, 群落资源消耗, 种内竞争加剧, 蒙古冰草种群分布格局发生改变。2) 种子的扩散限制。种子扩散是种群动态中极为重要的生态过程, 种子的扩散方式与种群的空间聚集强度和尺度有关[23, 24]。在草地生态系统中, 植被盖度是影响种子扩散的因素之一[25], 本研究中, 3个群落植被盖度差异明显, 植被盖度的变化可以改变风速, 加之植被盖度对种子的拦截作用, 因而影响了种子的传播距离和聚集程度[23]。3) 群落生境资源的异质性。不同植物群落具有其特有的生态性能与生境特征, 牛枝子是一种豆科多年生小半灌木, 根瘤菌不仅具有固氮作用, 而且对地表凋落物拦截聚集能力较强, 有利于促进土壤养分富集效应的形成[26]。杨阳等[27]的研究表明, 荒漠草原不同优势物种的根际土壤养分差异显著, 依次表现为豆科[苦豆子(Sophora alopecuroides)]> 禾本科[长芒草(Stipa bungeana)、蒙古冰草]> 萝藦科(牛心朴子)。也有研究表明, 短花针茅[13]与牛心朴子[28]群落所在生境一般较为贫瘠, 土壤含水量较低, 物种单一。因此, 在资源有限的情况下, 植物只能通过调节自身的形态、密度与分布策略来适应生境。相对蒙古冰草+牛枝子群落, 蒙古冰草种群在短花针茅+蒙古冰草与蒙古冰草+牛心朴子群落中的密度、盖度和地上生物量均较低, 聚集尺度与强度也较低, 可能正是这一原因所致。
群落在长期演替过程中, 各种群之间既相互依存又相互竞争, 种群的空间关联性能够反映这一复杂的竞争关系与分异规律[29]。荒漠草原区, 水分是限制植物生长、繁殖与分布的关键因子[20], 不同物种对资源的趋同需求势必会导致物种之间的竞争, 而植株密度和盖度的变化是决定植物种内种间关系的重要因子[22]。在蒙古冰草+牛枝子群落, 蒙古冰草种群的密度、盖度均大于其他两个群落, 间接表明该群落为蒙古冰草种群提供了较为良好的生存繁衍条件, 使其形成较大的植被斑块, 从而对牛枝子种群可利用资源的获取造成威胁, 使得二者之间表现出显著的竞争关系, 种间关联性在0~2.9 m尺度上表现为负相关; 随尺度增大, 种内竞争加剧, 而种间竞争减弱, 使种间关联性开始由负关联转变为无关联。蒙古冰草与短花针茅种群仅在0~1.0 m尺度上出现负关联, 表现为竞争的生态关系, 随尺度增加表现为无关联。蒙古冰草与短花针茅均属多年生禾草, 在长期干旱的气候条件下, 都演替成为宁夏荒漠草原的优势种群, 有其独立的分布空间, 群落结构稳定, 因此在同类资源条件下, 二者的竞争能力相当, 仅在小尺度范围内由于个体数量多, 生态位重叠较大而产生资源竞争[13, 30]; 随尺度增大, 个体密度减少, 种群间生态位重叠减弱, 种群个体表现为独立分布, 种群之间逐渐趋向无关联[22, 30]。牛心朴子根系发达, 繁殖系数高, 具有极强的生境适应能力, 在生态学上常被认为是草原逆行演替过程中的指示物种, 是干旱与半干旱草原、荒漠草原沙质荒漠化的标志[28]。本研究中牛心朴子种群与蒙古冰草种群在整个研究尺度内没有显著的相关性, 可能是物种的生境偏好不同, 使得两者之间的生态位分化明显, 种群间表现出无联性。荒漠草原生境的破碎化也会降低物种对资源利用的相似性, 加速物种间生态位分化[31]。
不同群落生境蒙古冰草种群的数量特征差异显著(P< 0.05), 在小于4 m尺度范围内各群落生境蒙古冰草种群主要表现为聚集分布, 随尺度增大, 聚集强度减弱, 逐渐过渡到随机分布和均匀分布。蒙古冰草种群与短花针茅种群、蒙古冰草种群与牛枝子种群在小于4 m尺度上呈显著负关联, 表现为明显的竞争关系, 随研究尺度增大趋于无关联。综上, 蒙古冰草种群在宁夏荒漠草原小尺度范围内主要以聚集的斑块形式存在。为提高幼苗竞争, 在退化荒漠草原的恢复与重建过程中, 适宜以较高密度进行补播。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|