越冬时葎草雌雄株生理反应和再生策略的差异分析
[何林江1  , 黄曦叶
, 黄曦叶1 , 刘金平1, * , 游明鸿2 , 杨小琴1 ]
, 黄曦叶, 游明鸿|
|


作者简介:何林江(1994-),男,四川梓潼人,在读硕士。E-mail: 872162384@qq.com
以野生雌雄葎草为材料,通过测定越冬前(12月1日)、越冬中(1月15日)和越冬后(4月15日)雌雄株构件中可溶性糖(SS)、淀粉(ST)、丙酮酸(PA)含量和硝酸还原酶(NR)活性,测定越冬后构件存活率和再生率及再生构件性状与生物量分配比,分析雌雄株越冬时生理反应和再生策略的差异。结果表明:1)越冬前雄株叶片中ST含量小于雌株而SS含量大于雌株,其成熟茎和根的SS和ST含量均小于雌株( P<0.05),雄株通过提高叶中SS含量抵御寒冷,雌株通过转移糖分到茎和根应对寒冷;2)越冬前雄株叶和茎的PA含量显著大于雌株,根中含量显著小于雌株( P<0.05),构件中NR活性无性别差异( P>0.05),雄株通过增强茎和叶的呼吸抵御寒冷,雌株通过根储备营养应对寒冷;3)越冬后雌株再生叶的SS和ST含量大于雄株( P<0.05),PA含量和NR活性小于雄株,其再生代谢基础高于雄株;4)越冬时雄株植株存活率为26.67%,为雌株植株存活率的34.78%,雄株存活植株的茎存活率仅为雌株的32.24%,越冬后雌株茎、叶再生率仅为雄株的21.03%和23.82%,而花序再生率高达74.40%;5)雌株再生叶数和叶面积及再生茎长小于雄株( P<0.05),其再生茎和叶生物量分配比仅0.59%和1.31%,再生花序分配达98.09%,其再生花序柄和花序轴长及小花数大于越冬前正常花序,且萼片面积增大且生物量分配比占31.26%,总面积达624.92 cm2·株-1,为再生叶总面积的24.50倍。越冬时雄株通过生理反应抵御寒冷,使茎和根贮藏物质低于雌株,故其存活率和再生率低于雌株。越冬后雄株再生生物量仅1.57 g·株-1全部分配到营养器官,雌株再生总生物量达32.05 g·株-1,且98.09%分配到花序,萼片代替叶片为主要光合器官,为花序形成和开花就近提供养分。
This research studied the wild dioecious climbing herb Humulus scandens, in order to analyze the differences in physiological response and regeneration strategies between male and female plants, while over-wintering. Various physiological data, including soluble sugar (SS), starch (ST), and pyruvic acid (PA) contents and nitrate reductase (NR) activity of male and female plant parts (young leaves, mature leaves, mature stems, roots) were determined before winter (December 1), during over-wintering (January 15) and after winter (April 15). Survival rate and regeneration rate of plant parts biomass allocation ratio were also determined after wintering, to give insight into differential biomass allocation strategy of male and female plants. The results showed: 1) The ST content in the leaves of male plants before wintering was lower than that in female plants, and the content of SS and ST in mature stems and roots were lower than those in female plants ( P<0.05). Male plants resisted cold by increasing SS in leaves, while female plants responded to cold by transferring sugar to the stems and roots. 2) The PA content of male plant leaves and stems was significantly higher than that in female plants before wintering, but PA content in male plant roots was significantly lower than that in female plants ( P<0.05). There was no gender difference in NR activity of plant modules ( P>0.05). Male plants resisted cold by enhancing the respiration of stems and leaves, while female plants responded to the cold through utilization of substrate reserves in the roots. 3) The content of SS and ST in the regenerated leaves of female plants was higher than in male plants ( P<0.05), and the PA content and NR activity in leaves were both lower than in male plants. The PA content increased over winter in the female mature stems yet decreased in male plants ( P<0.05). The metabolic activity involved in female plant regeneration was higher than that of male plants. 4) The survival rate of over-wintering male plants was 26.67%, while survival rate of female plants was 76.7%. The survival rate of stems in male plants was only 32.24% of that in female plants (12.50% and 38.77%, respectively). The regeneration rates of plant parts were, for male and female plants, respectively: leaves 5.85% and 1.23%, stems 13.10% and 3.12%, while inflorescence regeneration in female plants was 74.40%. 5) The number of regenerated leaves, the leaf area, and the length of the regenerated stems of female plants were all lower than those of male plants ( P<0.05). The total biomass proportion contributed by regenerated stems and leaves was only 0.59% and 1.31%, respectively, while the contribution to total biomass of regenerated inflorescences was 98.09%. The regenerated inflorescence stalk and axis length and floret number of regenerated inflorescences were larger than those of inflorescences observed before winter. Sepals exhibited increased area and greenness, and accounted for 31.26% of total biomass in regenerated plants, and their total area was as much as 625 cm2·plant-1, compared with a total area of regenerated leaves of 25.51 cm2·plant-1. During overwintering, male plants resisted cold through physiological response, in which the stored substrates of male stems and roots were lower than those of female plants. Thus the survival rate and regeneration rate of male plants were lower than those of female plants. After wintering, the biomass of regenerated male plants was only 1.57 g·plant-1 and all of this was allocated to vegetative organs. By contrast, the average biomass of female plants was 32.05 g·plant-1, and 98.09% of this was allocated to inflorescence formation with sepals instead of leaves being the main photosynthetic organs, providing nutrients for inflorescence formation and nearby flower development.
雌雄异株植物维持种群稳定性的能力较差[1], 雌雄个体生长、生理、生殖及抗性等有明显的性别差异[2], 生境胁迫易引起种群性比失衡而影响繁殖能力, 甚至造成物种的濒危或灭绝[3]。尤其对于雌雄异株草本攀援植物, 因集性别分化性、对支持物的依赖性和草本植物的短命性于一身[4], 故要求雌、雄株依据胁迫种类和强度, 快速做出匹配协同的应对策略, 顺利完成生活史, 才能保证物种的延续。
有药用、饲用、生态绿化、工业原料及水土保持材料等利用价值[5, 6]的葎草(Humulus scandens), 为桑科葎草属雌雄异株草本攀缘植物。雌、雄株在生长发育、构件性状、生殖投入等方面有显著差异[7, 8, 9], 雌、雄株对支持物、温度、水分、光照等生境因子表现出不同的生长策略和繁殖对策[10, 11, 12, 13], 凭借极强的形态可塑性和抗性系统, 通过调节生长、生理和生殖过程, 逃离、抵御和忍耐生境胁迫, 表现出极强的抗逆性能[4, 14]。经霜后葎草地上部分或全株死亡, 降温幅度和霜期长短决定植株能否存活与再生, 故亚热带湿润季风气候区, 葎草表现为一年生或多年生特点。诸多研究表明, 雄株具有更早的抗寒形成和更高的耐冻性[10, 15], 冷害对雄株叶绿体和膜结构伤害低于雌株, 雄株具有更高的耐寒性和抗寒性[1, 10, 16]。但野外调查发现, 越冬后葎草雌株存活率远大于雄株, 表明雌株抗寒力强于雄株。
抗寒性受植物种类、发育时期、生理状态、组织器官及低温程度及胁迫强度的影响。木本与草本植物生物学特点不同, 其生长周期、生长习性和形态结构差异, 越冬时物质流动方向和能量贮藏部位不同, 故两者抗寒形成和抗性结果可比性较小。且现有结果多是室内盆栽骤然降温下获得, 野外生境下植株根系深度大于盆栽, 植物对逐渐降温有逐步响应的过程, 开展自然降温下雌雄异株植物应对寒冷的性别差异研究, 更能真实反映雌雄个体抗寒性差异的机理, 对该类植物保护和利用更具有指导价值。
本试验以野生葎草为材料, 通过测定越冬前、越冬中、越冬后, 雌雄株可溶性糖(soluble sugar, SS)、淀粉(starch, ST)、丙酮酸(pyruvic acid, PA)含量和硝酸还原酶(nitrate reductase, NR)活性差异, 测定越冬后植株和构件存活率及构件再生率, 测定再生构件性状及生物量分配比, 研究越冬时葎草雌雄株生理反应和再生策略差异。拟回答以下问题:1)越冬时雌雄株糖分转移和呼吸强度是否有性别差异?2)雌雄株植株和构件存活率及构件再生率是否有性别差异?3)雌雄株再生构件性状及生物量分配是否有性别差异?分析雌雄葎草越冬策略和抗寒性差异, 旨在探讨葎草雌株抗寒性大于雄株的生理基础和再生对策, 以期为雌雄异株植物抗寒性研究奠定基础, 为雌雄异株草本攀缘植物资源的保护利用提供借鉴。
试验于四川省南充市顺庆区西华师范大学新校区进行, 为典型的亚热带湿润季风气候, 地理坐标为北纬30° 82'、东经106° 06', 年平均气温17.4 ℃, 最高气温40.1 ℃, 最低气温-2.8 ℃, 日照时间1266.7 h, 有霜期仅13.7 d, 年降水量1020.8 mm。土壤为紫色土, pH为6.6, 有效氮、有效磷、有效钾含量分别为29.41 mg· kg-1、8.13 mg· kg-1和74.06 mg· kg-1, 有机质含量0.89%。测定期间温度情况见表1。
| 表1 2017和2018年冬春季温度情况 Table 1 Winter and spring temperature in 2017 and 2018 (℃) |
2017年3月, 在平坦退化绿地中, 清理杂物, 对野生葎草幼苗进行逐次间苗, 最终形成均匀分布、密度约为4株· m-2葎草种群, 于6月随机选取健壮、大小相同雌株、雄株各30株为材料, 期间避免植株交互缠绕。
1.3.1 生理代谢 于越冬前(2017年12月15日)、越冬中(2018年1月15日)和越冬后(2018年4月15日), 随机选择雌雄株各3株, 分别取青绿的成熟叶和幼嫩叶各10片(没有绿叶则不测定), 取2 cm长的成熟茎和根各10段, 剪碎混合均匀备用。构件样品各称量0.2 g, 用苏州科铭生物有限公司生产的试剂盒, 分别测定与计算可溶性糖(SS)、淀粉(ST)、丙酮酸(PA)含量和硝酸还原酶(NR)活性[17], 3次重复。
1.3.2 存活率和再生情况 存活率:越冬前和越冬中, 随机选雌雄株各6株, 测单株的茎、叶、花序数。越冬后以新茎叶形成为植株存活依据, 随机选存活雌雄株各6株, 测存活构件数, 计算:植株存活率=存活植株数/越冬前植株数× 100%, 构件存活率=存活构件数/越冬前构件数× 100%, 构件再生率=再生构件数/越冬前构件数× 100%。
再生构件性状:越冬后选存活雌雄随机各6株, 测定叶片数、单叶面积和总叶面积, 分枝数和茎长, 花序数、花序柄(轴)长、小花数和萼片面积。
生物量分配:越冬后选存活雌雄随机各6株, 分离再生茎、叶、花, 称鲜重后, 分别装袋。于105 ℃烘至恒重, 称干重为生物量。计算:构件含水量=(构件鲜重-构件干重)/鲜重× 100%, 生物量分配比=构件生物量/总生物量× 100%。
用SAS 10.0软件对所有数据进行方差分析和析因分析, 并用Duncan法对各参数进行显著性检验(SNK检验)。
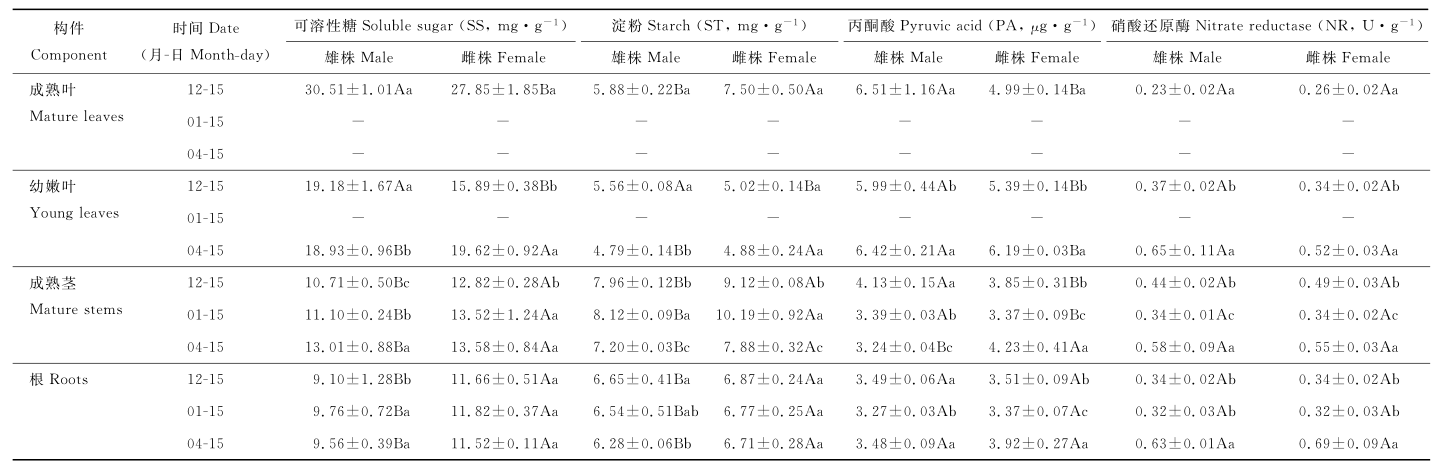
2.1.1 糖分 越冬前, 成熟叶、幼嫩叶、成熟茎和根的SS和ST含量有显著性别差异(P<0.05)(表2), 雄株成熟叶和幼嫩叶的SS含量显著高于雌株, 成熟叶ST含量显著低于雌株, 而幼嫩叶ST含量显著高于雌株(P<0.05)。越冬时雄、雌株叶片都凋谢死亡, 而越冬后雌株幼嫩叶的SS和ST含量大于雄株(P<0.05)。整个越冬过程中, 雌株成熟茎和根的SS和ST含量均显著大于雄株(P<0.05)。
| 表 2 雌雄株构件可溶性糖、淀粉、丙酮酸含量和硝酸还原酶活性的动态变化 Table 2 Dynamic changes of SS, ST, PA content and NR activity in male and female plants |
越冬前, 雄、雌株构件的SS含量为成熟叶> 幼嫩叶> 成熟茎> 根, ST含量为成熟茎> 根> 成熟叶> 幼嫩叶。越冬时随叶片死亡, 成熟茎中SS和ST含量显著增加(P<0.05), 雄株根的SS增加较多而ST增加较少, 雌株根的SS和ST含量变化较小。越冬后恢复生长时, 雄、雌株根的SS和ST含量均下降(P>0.05), 而成熟茎的ST含量显著降低, 雄株茎中SS含量增加大于雌株, 雌株再生叶的糖分含量显著高于雄株(P<0.05)。
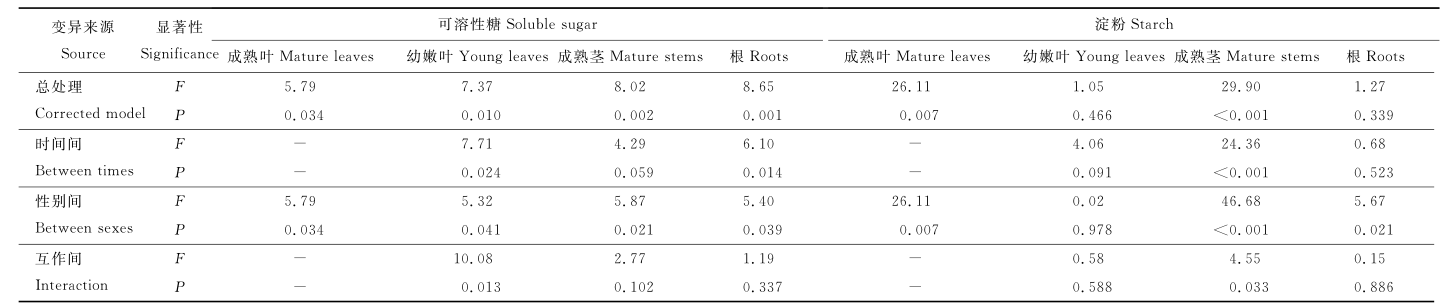
方差分析表明, 越冬过程中, 除幼叶ST含量外, 其他构件中SS和ST含量有显著的性别差异(P<0.05) (表3), 由F值可知, SS和ST性别差异为成熟茎> 成熟叶> 根> 幼嫩叶。越冬对幼嫩叶和根的SS含量有显著影响, 对成熟茎的ST含量有极显著影响(P<0.01)。性别对成熟叶、成熟茎中SS含量, 对成熟叶、成熟茎和根中ST含量影响大于时间, 越冬时间对幼嫩叶SS和ST含量及根的SS含量的影响大于性别。性别与时间对幼嫩叶SS和成熟茎ST含量有显著互作作用(P<0.05)。
| 表 3 雌雄株构件可溶性糖和淀粉含量的双因子方差分析 Table 3 Two-factor variance analysis about SS and ST content in components of male and female plants 。。 |
2.1.2 丙酮酸 越冬过程中, 成熟叶、幼嫩叶、成熟茎的PA含量有显著性别差异(P<0.05)(表2), 根的PA含量无性别差异(P>0.05)。越冬前, 雄株成熟叶、幼嫩叶、成熟茎的PA含量显著大于雌株, 根的含量小于雌株。越冬中, 雄、雌株成熟茎和根的PA含量显著降低(P<0.05)。越冬后, 雄、雌株根的PA含量显著增加, 雌株成熟茎的PA含量增加而雄株降低(P<0.05), 雄株再生幼嫩叶PA含量大于雌株。
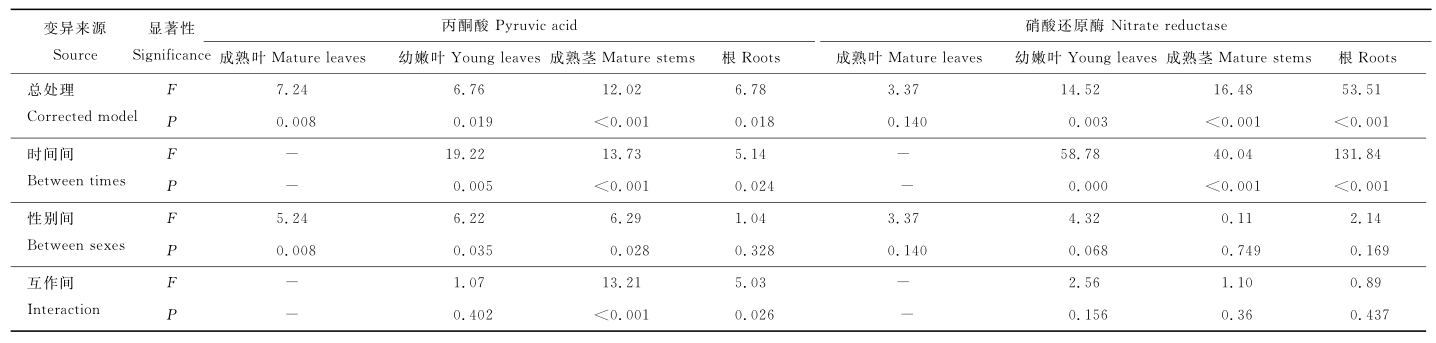
方差分析表明, 越冬过程中, 构件的PA含量均有显著变化(P<0.05) (表4), 总处理的影响为成熟茎> 成熟叶> 根> 幼嫩叶。性别对根的PA含量无影响, 时间是影响构件中PA含量的主要因子。性别和时间对成熟茎和根的PA含量有显著互作(P<0.05)。
| 表 4 雌雄株构件中丙酮酸含量和硝酸还原酶活性的双因子方差分析 Table 4 Two-factor variance analysis about PA and NR content in components of male and female plants |
2.1.3 硝酸还原酶 越冬过程中, 根、茎、叶的NR活性无性别差异(P>0.05)(表2)。越冬前, 成熟茎的NR活性最大, 成熟叶最小。越冬中, 成熟茎和根的NR活性下降为最小值。越冬后, 根、茎、叶的NR活性均显著增加(P<0.05), 根和叶增加幅度大于茎, 雄株茎、叶增加大于雌株, 根的增加小于雌株。
方差分析表明, 越冬过程中, 幼嫩叶、成熟茎和根的NR活性均有极显著变化(P<0.01)(表4), 总处理的影响为根> 成熟茎> 幼嫩叶> 成熟叶。性别对构件的NR活性无影响, 时间是影响构件中NR活性的主要因子。性别和时间对NR活性无显著互作。
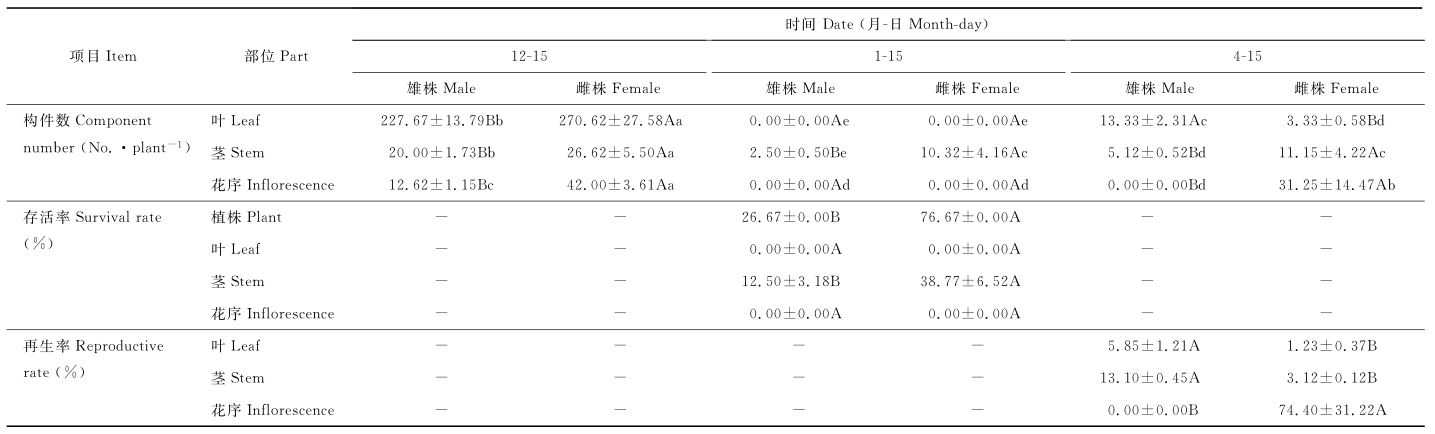
2.2.1 存活率 越冬前, 雌株茎、叶、花序数均大于雄株(P<0.05)(表5), 越冬时雄、雌株的叶和花序全部凋谢干枯, 茎全部干枯造成植物死亡, 部分茎死亡则植株可以存活再生。越冬时雄、雌株都有植株死亡, 植株存活率有显著性别差异(P<0.05), 雄株存活率显著低于雌株, 仅为雌株的34.79%。存活植株上的茎存活率有显著性别差异(P<0.05), 雌株茎存活率显著高于雄株, 雄株茎存活率仅为雌株的32.24%。方差分析表明, 越冬过程中, 雄、雌株的植株存活率和茎存活率有极显著差异(P<0.01)(表6), 性别是决定植株存活率和茎存活率的主要因素, 性别对植株存活率的影响大于茎存活率。
| 表 5 雌雄株构件存活率和再生率的动态变化 Table 5 Dynamic changes of components survival rate and reproductive rate of of male and female plants |
| 表6 构件存活率和再生率的双因子方差分析 Table 6 Two-factor variance analysis about survival rate and reproductive rate of components |
2.2.2 再生率 存活植株的茎、叶、花序数再生率有显著性别差异(P<0.05)(表5), 雄株的叶和茎再生率显著大于雌株, 越冬时雌株成熟茎存活率大于雄株, 但以成熟茎为基础的茎、叶再生率仅为雄株的23.82%和21.03%。越冬时雄、雌株花序全部凋谢干枯, 越冬后雄株花序未再生, 而雌株花序再生率高达74.40%。方差分析表明, 性别对茎、叶、花序数再生率有极显著差异(P<0.01)(表6), 对花序再生率的影响最大, 对叶再生率次之, 对茎较小。
2.3.1 再生构件性状 雄株再生叶的单叶面积显著小于雌株(P<0.05)(表7), 因雄株再生叶片数是雌株的4.76倍(表5), 使雄株总叶面积显著大于雌株, 是雌株的2.26倍。越冬时雌株茎存活率是雄株的3.10倍, 虽其茎再生率仅为雄株的23.82%, 越冬后雌株茎数显著大于雄株(P<0.05) (表5), 但其再生茎的茎长显著低于雄株(P<0.05) (表7), 仅为雄株再生茎长的26.51%。
| 表7 雌雄株再生构件性状和生物量分配比 Table 7 Characters of regeneration components and biomass distribution ratio of male and female plants |
越冬前雌株花序数和越冬后再生花序数显著大于雄株(P< 0.05)(表5), 其再生花序的花序柄和花序轴长及小花数显著增加(越冬前花序柄长为1.14 cm, 轴长为2.34 cm, 小花数为6.60), 萼片面积增大且绿化(越冬前花序的单萼面积为0.22 cm2), 使雌株的萼片总面积达624.92 cm2(表7), 是总叶面积的24.50倍。
2.3.2 生物量分配 雄株再生茎、叶生物量和分配比显著大于雌株(P< 0.05)(表7), 雄株再生茎、叶生物量是雌株的3.58和2.12倍, 分配比是雌株的73.41和43.27倍, 但再生总生物量仅1.57 g· 株-1。雌株再生总生物量达32.05 g· 株-1, 其中花序生物量为31.44 g· 株-1, 分配比达98.09%, 其中萼片分配比占31.26%。
植物通过调整自由水和束缚水比值降低植株含水量[18], 调节酶的活性和气孔开合度降低呼吸速率[19], 改变激素组成与比例抑制生长而进入休眠状态, 水解淀粉降低冰点增加保护性物质等复杂的生理代谢途径, 提高植物的抗寒性为安全越冬提供保障。葎草具有较强的抗寒性, 通过降低光合速率、限制SS合成, 促进呼吸作用、加速SS分解来应对低温[20]。本试验中, 雌雄株采取不同的生理代谢方式为越冬做准备, 雄株通过提高叶中SS和PA含量抵御寒冷, 雌株通过提高茎和根ST含量应对寒冷。叶片中SS可调节细胞液浓度避免蛋白质胶体遇冷变性凝聚, 也可转化为磷脂、氨基酸等保护物质和能源, 以维持细胞膜的完整性[21], 雄株叶中消耗大量SS提高呼吸作用第一阶段产物PA含量的同时, SS含量显著高于雌株, 但此时雄株叶的叶绿素下降速度大于雌株, 且现实和潜在光合速率低于雌株[22], 其SS是由其他物质转化或其他构件转移而来, 故其成熟茎和根的SS、ST含量均小于雌株。霜害或冻害前, 雌雄株的叶片数无性别差异, 或许与寒冷引起叶片水势增加和根系吸水降低等水分代谢的性别差异有关。葎草雄株体内含水量小于雌株[23], 雄株对水分缺乏更加敏感, 且优先把更多水分分配到叶片[24], 寒冷下雄株更易降低水势引起植株失水, 是否直接影响SS的运转和累积及呼吸强度, 待于深入研究。
PA是糖代谢及体内多种物质相互转化的重要中间体, NR是调控氮代谢的关键酶[17]。越冬前雄株主要通过提高呼吸作用抵御寒冷, 雌株主要通过提高贮备物质应对寒冷, 故雄株叶和茎中PA含量大于雌株, 而根中PA和ST含量小于雌株。越冬中雌雄株PA含量和NR活性均降为最低值。越冬后茎和叶的PA含量有显著性别差异, 而性别对根的PA含量无影响, 但整个越冬过程中各构件的NR活性无性别差异, 表明糖代谢途径及物质转化效率是决定雌雄株生理差异的主要因素, 性别对氮代谢影响较小。氮代谢与糖代谢密切相关, 25%糖代谢能量用于氮代谢[25], 某些组织中氮代谢可消耗掉55%的糖代谢能量, 氮代谢易受生境条件及构件部位的影响[26]。本试验仅测定了越冬前后构件中PA含量和NR活性, 对于糖代谢和碳代谢速率变化及协同关系对抗冻性影响的性别差异尚缺, 同一性别植株死亡与存活的物质代谢有何差异, 同一植株死亡茎和存活茎的结构构造和代谢能力有何差异, 待于系统研究。
葎草茎内木质部不发达, 主要由膨压支撑植株生长, 寒冷使雄株叶片水势和气孔导度增大率、叶绿素含量和光合速率降低率大于雌株[22], 使越冬前雄株茎、叶和花序数低于雌株。因雌、雄株采取不同的生理代谢和物质转移途径抵御或应对寒冷, 虽霜害或冻害使体内的液体结成冰晶, 原生质严重脱水或机械损伤, 造成雌雄株叶片与花序全部干枯凋谢, 但雌株茎的抵御冻害能力高于雄株, 致使雌株的植株存活率和茎存活率显著高于雄株。葎草无茎基分蘖能力, 仅通过分枝进行分生, 即使根具有活力也不会再生为完整植株, 茎存活数量是决定植株死亡与否及再生能力的关键。茎的抗冻性受发育阶段、生理状态、低温程度和持续时间等影响, 本试验中, 越健壮、饱满、木质化茎的存活率越高, 该冬季气温低于0 ℃仅3 d, 最低温为-1 ℃, 葎草存活率较高, 但诸多年份雌雄株全部死亡, 关于茎发育性状与低温胁迫强度对抗冻性影响的性别差异, 待进一步深入研究。性别是决定植株和茎存活率的主要因素, 支持物对分枝率、枝条密度、茎长及茎径影响的性别差异[2, 4], 也会导致雌雄株的越冬率不同。野外调查发现, 匍匐于地表的种群密度越大, 位于底层茎的越冬率越高。攀援于灌木上的裸露枝条全部死亡, 而攀援于乔木且受树冠遮阴的葎草, 雌、雄株全部存活且叶未完全凋谢。支持物对气温影响极小, 支持物形态主要决定霜降于叶片的难易度和受霜害叶片的比例, 早霜发生时间和降温幅度是影响葎草越冬率的外因, 对冬季降温的生理响应及物质转化或转移效率是决定越冬率的内因。
构件性状可反映植物对生境的适应策略, 也可反映生境对植物生长的影响程度[27]。植株通过功能权衡与物质分配, 形成适应生境的构件形态和生理特征[28]。目前对胁迫生境下葎草生长、生理、生殖和抗性策略进行了诸多研究, 表明雌雄株均能通过构件性状和异速生长的形态可塑性, 形成适应、逃离或抵御生境胁迫的生长与繁殖策略[1, 14, 20, 24]。但越冬后雄株再生叶数和叶面积及再生茎长显著大于雌株, 利于雄株营养生长和拓展空间, 而雌株主要进行生殖再生, 形成比常规生殖更多更长的花序数和小花数, 构件数量异化和生物量结构失衡表现出雌雄株截然不同的再生策略。性别分化使雌、雄株在种群形成和物种延续中任务不同, 通常雄株把更多的物质和能量用于生殖分配, 利于提供足量的花粉, 雌株生殖分配相对稳定且根分配较高为孕育种子和提高抗性提供保证[14, 24, 29]。越冬后雄株放弃生殖再生, 雌株几近放弃再生茎叶, 而由花序上膨大的萼片代替叶片为主要光合器官, 就近满足花序的营养需求, 雌株再生出数倍于常规花序的花序柄、花序轴和小花数, 便于吸引传粉昆虫和接受种群外花粉。虽葎草对光周期不敏感, 开花无须春化作用[30], 如何按有效积温法则和花芽分化原理及物质分配机理, 解释越冬后雌雄株再生策略差异的机理, 待于深入研究。
1) 越冬前雄株通过提高叶片SS和PA含量, 增加茎和叶的呼吸强度抵御寒冷, 雌株通过贮藏ST到茎和根应对寒冷。
2) 越冬时雌株抗冻生理代谢强于雄株, 越冬后雌株通过分解成熟茎中贮备物质为再生提供营养, 雌株再生的生理代谢基础高于雄株。
3) 越冬时雌雄株叶片和花序全部凋谢, 雄株的植株存活率为雌株的34.79%, 雄株存活植株的茎存活率为雌株的32.24%。
4) 越冬后雄株再生生物量为1.57 g· 株-1, 雌株为32.05 g· 株-1, 雄株仅进行茎和叶再生, 雌株98.09%再生生物量用于花序再生, 雌株茎、叶再生率仅为雄株的27.94%和47.19%。
5) 雌株再生叶数和叶面积及再生茎长小于雄株, 而再生花序的柄和轴长及小花数均大于正常花序, 且膨大叶化萼片为再生叶总面积的24.50倍, 萼片为雌株主要光合器官, 为开花结实就近提供营养。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|