




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
新疆半干旱区不同豆科/禾本科牧草混播草地生产力的变化研究
[谢开云1  , 曹凯
, 曹凯1 , 万江春1 , 王玉祥1 , 赵云2 , 朱进忠1, * ]
, 曹凯]
|
|


作者简介:谢开云(1984-),男,甘肃武威人,副教授,博士。E-mail: xkycah@163.com
混播草地竞争和促进关系是影响生产力和稳定性的重要因素,最终都通过牧草干物质产量以及草地物种的组成变化等指标表现出来。研究选择当地生产最常用的4种牧草无芒雀麦、鸭茅、红豆草、红三叶,以单播为对照,通过对2种豆/禾和3种牧草组合的混播草地牧草产量及物种组成变化进行长期监测,分析混播草地生产力的变化以及影响其稳定性的因素。结果表明:不论是单播还是混播,刈割干物质产量在年际间均表现为2017年高于2018年。放牧干物质产量在年际间均表现为2018年显著高于2017和2016年,鸭茅/红三叶混播草地除外。对2种牧草混播草地各物种干物质产量、密度及粗蛋白贡献的比例分析,从2016年到2018年,无芒雀麦干物质产量和密度所占比例均显著增加,鸭茅和红三叶干物质产量和密度所占比例均显著下降。红豆草与无芒雀麦混播时逐步降低,而与鸭茅混播时显著增加。3种牧草混播草地中,无芒雀麦和红豆草干物质产量和密度所占的比例显著增加,红三叶和鸭茅干物质产量和密度所占比例显著降低。综上所述,干旱区混播草地年际间干物质产量的消长变化主要取决于年均降水量和牧草的适应性。从混播草地干物质产量和密度的变化来看,随着年限增加,无芒雀麦和红豆草在混播草地中逐渐占据优势,鸭茅和红三叶在混播草地中逐渐处于劣势,且有消退的趋势。
The inter-species competitive and mutualistic relationships in mixed-species grassland are important to grassland productivity and stability. Ultimately, the cumulative effect of such factors is expressed by indicators such as yield and change in botanical composition in mixed-species grassland. In this study, monocultures of four most commonly sown forage species, smooth bromegrass ( Bromus inermis), orchard grass ( Dactylis glomerata), sainfoin ( Onobrychis viciifolia) and red clover ( Trifolium pratense), were established as controls, and the forage yields and changes in species composition of two-species combinations (one legume and one grass) and three-species combinations were monitored for three years, and changes in productivity and factors affecting sward stability were analysed. For both monoculture and mixed-species swards, the biomass as hay was higher in 2017 than in 2018. The grazing biomass yields of mixed-species grassland were also significantly higher in 2018 than in 2017 and 2016, except for mixed-species swards of orchard grass and red clover. From 2016 to 2018, the proportion of biomass and density of smooth bromegrass increased significantly, and the proportion of biomass and density of orchard grass and red clover decreased significantly. The proportion of biomass and density of sainfoin gradually increased when sown with smooth bromegrass, but it gradually decreased when sown with orchard grass. In mixed-species swards of three forages, the proportions of biomass and density of smooth bromegrass and sainfoin increased significantly, while those of red clover and orchard grass decreased significantly. In summary, the variation of biomass over three years in mixed-species grassland in the arid conditions of this experiment was found to depend mainly on the annual average rainfall and the adaptability of the species present. From the changes in biomass and density observed in mixed-species swards, over time, smooth bromegrass and sainfoin gradually dominated while orchard grass and red clover were gradually lost and may disappear entirely in the longer term when included in mixed-species sowings in this region.
混播草地是人工草地建植的主要方式之一, 其中豆禾混播草地最为常见。豆禾混播草地中各种牧草能利用其生物学特性, 充分利用环境资源来提高牧草的产量和品质[1]。众多研究表明, 与单播草地相比, 豆禾混播草地牧草产量更高[2], 营养价值更均衡[3], 且能显著降低杂草发生率[4]。国内外已对豆禾混播草地进行了大量的研究, 主要集中在混播草地的牧草产量[5, 6]、营养价值[7, 8]、改善土壤[9, 10]、种间关系[11, 12]、群落稳定性[13]等方面。在混播草地中豆禾牧草之间存在着对水分、养分、空间、光照等资源的竞争, 最终都通过牧草生产力以及草地物种的组成变化等指标表现出来[14]。在豆禾混播草地中不仅存在着竞争关系, 也存在着促进关系[15], 即豆科植物固定大气中的氮不仅可以满足自身的生长需求, 还可以通过各种途径为伴生的禾本科植物提供氮源[16, 17]。这两种关系的平衡是影响其高产稳产的重要因素之一。建植的人工混播草地能够发挥出预期生产力的前提是维持群落的稳定性, 而选择正确的品种组合、环境因子的合理调控、管理和利用的适度性是维系人工草地群落稳定性的根本保证[18]。因此, 建植多年生人工混播草地不仅要考虑光照、温度、水分等环境因素, 还要考虑牧草生物学特性、管理措施、利用方式等因素。
新疆作为我国主要牧区之一, 由于不合理利用和全球气候变化等因素, 导致天然草地生产力下降, 生态环境极度恶化[19, 20]。在实施草场休牧、禁牧的新政策环境下, 畜牧业的重心也开始由草原畜牧业向农区舍饲畜牧业转移, 然而饲草料短缺一直是制约新疆畜牧业现代化、规模化发展的重要因素[21], 而优质牧草生产对于解决当前新疆畜牧业发展中优质饲草料短缺的难题具有重要意义。在大力推进“ 种-养” 结合的前提下, 在新疆雨养山区建设优质高产的混播草地, 有布局地发展具有区域特色的生态畜牧业是响应国家号召, 实施“ 粮改饲” 发展农区草食畜牧业的重要战略布局。本研究通过长期观测不同牧草种类混播草地牧草产量和密度的变化来分析影响干旱半干旱区混播草地生产力的变化及影响因素, 以此来确定适宜该地区混播草地的利用方式和管理措施, 这将为该地区生产牧草、发展草食畜牧业以及生态系统的恢复提供重要的理论和科学依据。
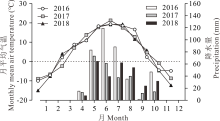
大田试验于2016、2017和2018年在新疆乌鲁木齐市谢家沟地区(43° 31'4″ N, 87° 03'4″ E, 海拔1675 m)开展。研究区地处天山北坡中、低山带的山溪坡阶地上, 属典型中温带大陆性气候。年均气温2.1~3.3 ℃, 年均降水量100~400 mm, 降水季节性分布不均, 主要集中在4-10月。年蒸发量为1141.7~1283.3 mm, 生长期短, 无霜期仅113~130 d。参照前人指标以年降水量≤ 200 mm为干旱区, 200~400 mm为半干旱区[22], 并结合2016、2017和2018年实测降水量, 分别为389.6、250.1和282.5 mm, 该地区被定义为半干旱区(图1)。试验区建植人工草地前是退化草地, 主要建群植物为蓝花棘豆(Oxytropis coerulea)、西伯利亚蓼(Polygonum sibiricum)、鹅绒委陵菜(Potentilla anserina)、蒲公英(Taraxacum mongolicum)等, 鼠害严重, 牧用价值已近丧失。土壤为沙质土, 含有较多的砂砾质, 肥力较低, 试验地0~30 cm土层土壤基础理化性状参数列于表1。
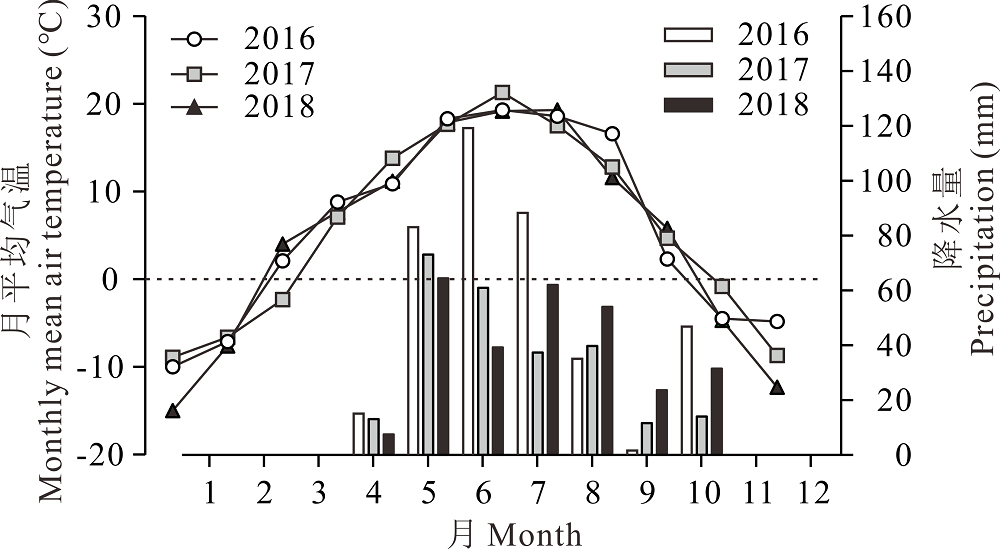 | 图1 试验期间试验地月平均气温和降水量Fig.1 Precipitation distribution and air temperature in the experimental field during 2016 to 2018 |
| 表1 试验地0~30 cm土壤理化性质 Table 1 The properties of soil physical and chemical in 0-30 cm |
本实验采用随机区组设计, 以无芒雀麦(Bromus inermis)、鸭茅(Dactylis glomerate)、红豆草(Onobrychis viciaefolia)、红三叶(Trifolium pratense) 4种牧草单播为对照, 设置无芒雀麦/红豆草, 无芒雀麦/红三叶, 鸭茅/红豆草, 鸭茅/红三叶, 红三叶/无芒雀麦/红豆草, 无芒雀麦/红豆草/鸭茅混播。共10个处理, 每个处理重复4次, 每个小区15 m2(3 m× 5 m)。无芒雀麦、鸭茅、红豆草、红三叶的理论播种量分别为:37.5、22.5、30.0和22.5 kg· hm-2。2种牧草单播时, 各占1/2, 3种牧草混播时, 各占1/3。实际播种量根据各个牧草实际发芽率计算, 4种牧草的实际发芽率分别为84%、61%、50%、86%。选择这些物种, 一方面是因为它们在当地生产中最常用, 另一方面是因为它们有不同的形态特征(例如株高、叶型、分枝/分蘖数、根系构型等), 这些特征可能会使得它们对资源利用有不同的响应方式[23]。试验区于2016年6月8日播种, 播种方式为均匀撒播。播种后出苗期间进行杂草控制, 整个试验期间无灌溉。
1)牧草干物质产量测定:试验期间分别于2017年7月1日和2018年6月28日对所有小区进行刈割, 割草前测定各小区牧草产量, 记为第1茬牧草产量(刈割利用牧草产量); 割草后进行施肥, 所用肥料为磷酸二铵120 kg· hm-2(P2O5 46%)和硫酸钾100 kg· hm-2(K2O 50%)。从2016、2017和2018年9月30日-10月30日期间进行放牧, 放牧家畜为绵羊和肉牛混合放牧, 在放牧开始之前测定混播草地的牧草产量(测定日期分别为2016年9月26日、2017年9月28日和2018年9月22日), 记为第2茬牧草产量(放牧利用牧草产量)。小区干物质产量测定方法:所有小区随机选取1 m× 1 m的样方框内齐地面刈割后, 分种装纸袋称其鲜重, 并在65 ℃烘箱内烘至恒重, 用于计算其牧草干物质产量。混播中牧草产量占比=混播中各牧草产量/混播草地总牧草产量× 100%。
2)单位面积密度测定:为了监测牧草单位面积密度的变化, 每年测定牧草第2茬生物量时在所有小区1 m× 1 m的样方框内分种计量每种牧草的分枝/分蘖数(个)。混播中牧草密度占比=混播中各牧草密度/混播草地牧草总密度× 100%。
3)粗蛋白产量测定:分别将2016、2017和2018年第2茬牧草的烘干样品, 用植物粉碎机(FZ102)粉碎后用于测定牧草含氮量(采用凯氏定氮法), 测定方法参照鲍士旦[24]的土壤农化分析(第三版)。混播草地的粗蛋白产量=草地产量(kg· hm-2)× 牧草氮含量(%)× 6.25。
采用Excel 2013进行所有数据整理和汇总, 采用Graph Pad Prism 7.0软件制图, SAS 9.0 软件统计分析。用PROC GLM程序对刈割和放牧干物质产量、单位面积密度和粗蛋白产量进行方差分析。
对刈割干物质产量和放牧干物质产量进行方差分析, 结果表明, 混播草地的刈割(2017和2018年)和放牧干物质产量(2016、2017和2018年)年际之间具有极显著差异(P< 0.0001)。刈割和放牧干物质产量在不同混播方式之间也具有极显著差异(P< 0.0001)。但年份和混播方式之间的交互效应并不显著(P> 0.05, 表2)。
| 表2 刈割和放牧牧草干物质产量、单位面积密度和粗蛋白产量的方差分析 Table 2 Variance analysis result of forage dry matter yield (DMY) as hay and grazing, density and crude protein yield |
2017和2018年4种牧草单播的平均干物质产量分别为4361.77和2383.32 kg· hm-2, 两种牧草混播的平均干物质产量分别为5048.55和3623.36 kg· hm-2, 3种牧草混播的平均干物质产量分别为7239.95和4793.74 kg· hm-2。4种牧草单播(红豆草、红三叶、无芒雀麦和鸭茅), 2种牧草混播(无芒雀麦/红豆草、无芒雀麦/红三叶、鸭茅/红豆草、鸭茅/红三叶), 3种牧草混播(无芒雀麦/红豆草/红三叶、无芒雀麦/红豆草/鸭茅)的刈割干物质产量均表现出2017年高于2018年, 其中红三叶和无芒雀麦单播、无芒雀麦/红三叶、鸭茅/红三叶混播达到显著水平(P< 0.05, 图2)。
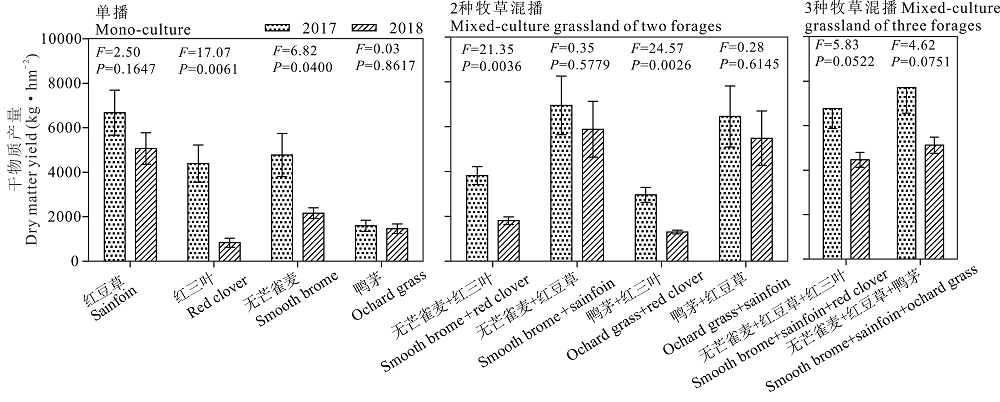 | 图2 不同混播处理下不同年份之间刈割利用牧草干物质产量的比较(第1茬)Fig.2 Comparison of dry matter yield as hay between different 2017 and 2018 under different planting mode (the 1st cutting) |
对于红豆草和无芒雀麦单播草地, 2018年放牧利用的干物质产量极显著高于2017和2016年(P< 0.001, 图3A), 对红三叶和鸭茅单播草地, 2018年放牧利用的干物质产量略高于2017和2016年, 但未达到显著水平(P> 0.05, 图3A)。对于2种牧草的混播草地, 无芒雀麦/红豆草和鸭茅/红豆草混播草地2018年放牧利用的干物质产量极显著高于2017和2016年(P< 0.01, 图3B), 无芒雀麦/红三叶混播草地干物质产量显著高于2017和2016年(P< 0.05, 图3B), 鸭茅/红三叶混播草地的干物质产量在2016、2017和2018年无显著差异(P> 0.05, 图3B)。对3种牧草混播草地, 无芒雀麦/红豆草/红三叶和无芒雀麦/红豆草/鸭茅混播草地均在2018年放牧利用的干物质产量极显著高于2017和2016年(P< 0.01, 图3C)。
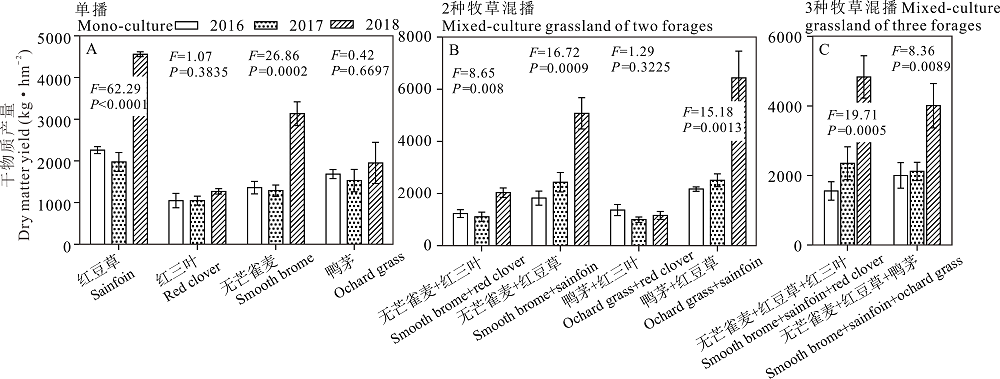 | 图3 不同混播处理下不同年份之间放牧利用牧草干物质产量的比较(第2茬)Fig.3 Comparison of dry matter yield as grazing among 2016, 2017 and 2018 under different planting mode (the 2nd cutting) |
随着年限的增加, 单播的红豆草和红三叶单位面积密度在逐渐降低, 单播的无芒雀麦单位面积密度先降低后极显著增加(P< 0.01, 图4A), 单播的鸭茅密度变化不明显。在混播中, 鸭茅/红三叶处理单位面积密度极显著降低(P< 0.01, 图4B)。红三叶/无芒雀麦/红豆草混播处理单位面积密度显著降低(P< 0.05, 图4C), 无芒雀麦/红豆草/鸭茅混播处理单位面积密度无明显的变化。
 | 图4 不同混播处理下不同年份之间放牧利用牧草单位面积分枝(分蘖)密度的比较Fig.4 Comparison of number of plant branches or tillers as grazing among 2016, 2017 and 2018 under different planting mode (the 2nd cutting) |
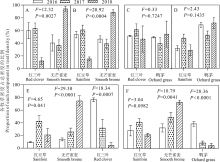
混播中各物种干物质产量所占比例不仅反映各物种对混播草地生产力的贡献率, 还能间接反映出各物种对环境资源的竞争能力。在红三叶/无芒雀麦混播草地中, 红三叶产量所占比例与2016(62.8%)和2017年(51.6%)相比, 2018年(6.4%)显著地降低(P< 0.05, 图5A), 相反无芒雀麦产量所占比例2018年(93.6%)比2016(37.2%)和2017年(48.4%)显著增加(P< 0.05, 图5A)。在红豆草/无芒雀麦混播草地中, 红豆草干物质产量所占比例从2016年到2018年逐步下降, 而无芒雀麦干物质产量所占比例从2016年到2018年逐步增加(P> 0.05, 图5B)。在红三叶/鸭茅的混播草地中, 红三叶和鸭茅的干物质产量所占比例随着年度的增加, 没有表现出明显的变化规律(P> 0.05, 图5C)。在红豆草/鸭茅的混播草地中, 红豆草干物质产量所占比例从2016年到2018年逐步增加, 而鸭茅干物质产量所占比例逐步下降(P> 0.05, 图5D)。
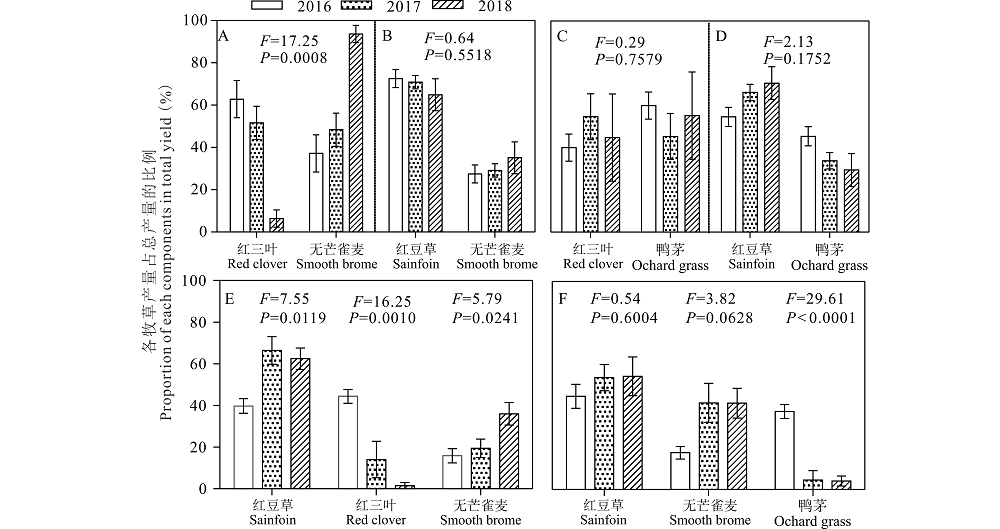 | 图5 不同混播处理下不同年份各种牧草干物质产量所占比例 A, B, C, D: 2种牧草混播处理Mixed-culture grassland of two forages; E, F: 3种牧草混播处理Mixed-culture grassland of three forages. 图6, 图8同The same with Fig.6 and Fig.8.Fig.5 Proportion of each components in total yield among 2016, 2017 and 2018 under different mixed treatments |
从2016年到2018年, 在红豆草/红三叶/无芒雀麦混播草地中, 红豆草干物质产量所占的比例显著增加(P< 0.05, 图5E)、红三叶干物质产量所占比例显著降低(从2016年的44.39%降低到2018年的1.53%, P< 0.05, 图5E), 无芒雀麦干物质产量所占比例显著增加(P< 0.05, 图5E)。在红豆草/无芒雀麦/鸭茅混播草地中, 红豆草干物质产量所占比例逐步增加(P> 0.05, 图5F), 无芒雀麦干物质产量所占比例逐步增加(P> 0.05, 图5F), 鸭茅干物质产量所占比例显著降低(从2016年的40%降低到2018年的5%以下, P< 0.05, 图5F)。
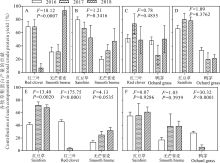
分别对2016、2017和2018年混播中各成分密度所占总密度的比例进行了分析, 结果表明, 在无芒雀麦分别与红三叶和红豆草混播中, 红三叶和红豆草密度所占比例在2016和2017年差异不显著, 2018年显著降低, 其中红三叶从59.69%和60.00%下降到12.08%(P< 0.05, 图6A); 红豆草从53.55%和60.97%下降到15.19%(P< 0.05, 图6B)。而无芒雀麦密度所占比例在2016和2017年无显著差异, 2018年显著增加, 与红三叶混播时从40.31%和36.63%增加到93.96%(P< 0.05, 图6A); 与红豆草混播时从46.45%和39.03%增加到88.61%(P< 0.05, 图6B)。鸭茅分别与红三叶和红豆草混播时, 鸭茅、红三叶和红豆草密度所占比例无明显的消长规律(P> 0.05, 图6C, D)。
在无芒雀麦/红豆草/红三叶和无芒雀麦/红豆草/鸭茅3种牧草混播中, 无芒雀麦密度所占比例显著增加(P< 0.05, 图6E, F), 红豆草密度所占比例先增加后减小(P< 0.05, 图6E), 而红三叶和鸭茅密度所占比例均显著下降(P< 0.05, 图6E, F)。这说明从混播草地密度的变化来看, 随着年限增加, 无芒雀麦在混播草地中逐渐占据优势, 鸭茅和红三叶在混播草地中逐渐处于劣势, 且有消退的趋势。
 | 图6 不同混播处理中不同年份各种牧草单位面积密度所占比例Fig.6 Proportion of forage components in total density among 2016, 2017 and 2018 under different mixed treatments |
对建植混播草地放牧利用的牧草(第2茬)粗蛋白产量进行分析, 结果表明, 对于4种单播牧草, 鸭茅的粗蛋白产量在2016、2017和2018年度之间差异不显著(P> 0.05)。红三叶、红豆草和无芒雀麦在2016和2018年之间差异不显著, 在2017年最高(P< 0.05, 图7A), 主要是因为2018年其干物质产量显著的增加(P< 0.05, 图3)。对于两种牧草混播草地, 无芒雀麦/红三叶混播草地在2017年粗蛋白显著大于2016和2018年(P< 0.05); 无芒雀麦/红豆草在2018年显著大于2016和2017年(P< 0.05); 鸭茅/红三叶在2016和2018年显著大于2017年(P< 0.05); 鸭茅/红豆草在2018年显著大于2016年, 2016年显著大于2017年(P< 0.05, 图7B)。对3种牧草混播草地, 红三叶/无芒雀麦/红豆草和鸭茅/无芒雀麦/红豆草混播草地粗蛋白产量在2018年显著大于2016和2017年(P< 0.05, 图7C)。
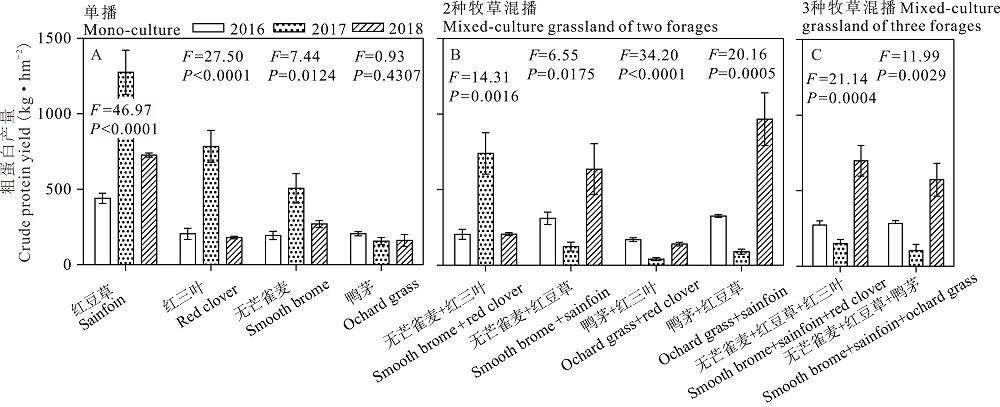 | 图7 不同混播处理下不同年份放牧草地粗蛋白产量比较Fig.7 Comparison of crude protein yield as grazing among 2016, 2017 and 2018 under different planting mode (the 2nd cutting) |
在红三叶/无芒雀麦的混播草地粗蛋白产量中, 2016和2017年红三叶粗蛋白产量占65%以上, 无芒雀麦的粗蛋白产量只占35%以下, 2018年红三叶的粗蛋白比例显著下降到10%以下, 而无芒雀麦的粗蛋白产量显著上升到90%以上(P< 0.05, 图8A)。在红豆草/无芒雀麦的混播草地粗蛋白产量中, 2016年红豆草的粗蛋白产量占80%, 无芒雀麦的粗蛋白产量只占20%, 随着年份增加, 到2018年红豆草的粗蛋白产量比下降到50%, 而无芒雀麦的粗蛋白产量比上升到50%(P> 0.05, 图8B)。在鸭茅/红三叶的混播草地粗蛋白产量中, 2016、2017和2018年红三叶和鸭茅粗蛋白比例变化无明显的规律(P> 0.05, 图8C)。在红豆草/鸭茅的混播草地粗蛋白产量中, 红豆草粗蛋白产量与2016年相比, 2018年增加了15%, 而鸭茅的粗蛋白产量比2016年降低了15%(P> 0.05, 图8D)。
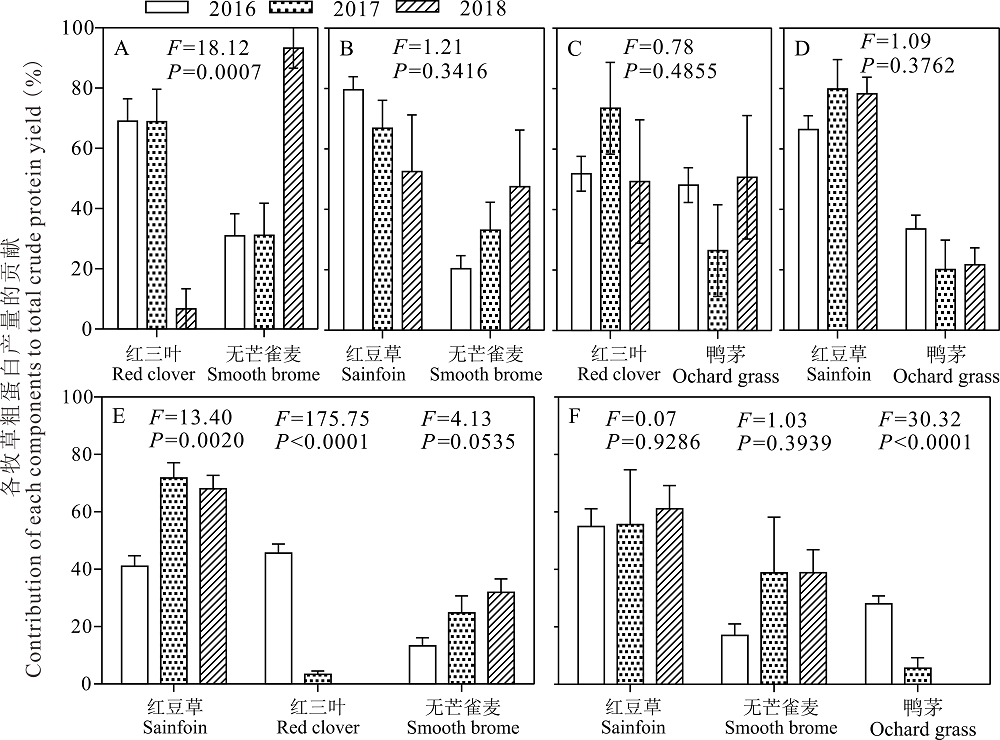 | 图8 不同混播处理中不同年份各种牧草粗蛋白贡献比Fig.8 Contribution of each components to total crude protein yield among 2016, 2017 and 2018 in different mixed treatments |
在红豆草/红三叶/无芒雀麦混播草地的粗蛋白产量中, 红豆草粗蛋白产量比例在2017(71.8%)和2018年(68.0%)比2016年(41.0%)显著增加(P< 0.05), 无芒雀麦粗蛋白产量比例在2017(24.8%)和2018年(31.9%)比2016年(13.3%)也有所增加(P> 0.05, 图8E), 相反红三叶粗蛋白比例从2016年的45.7%下降到2017年的3.4%, 2018年为0。
在红豆草/鸭茅/无芒雀麦混播草地的粗蛋白产量中, 红豆草的粗蛋白产量比例在2016、2017和2018年无显著变化(P> 0.05, 图8F), 无芒雀麦的粗蛋白产量2017和2018年与2016年相比有所增加(P> 0.05, 图8F), 相反, 鸭茅的粗蛋白比例从2016年的28.0%下降到2017年的5.6%, 2018年为0(P< 0.05, 图8F)。
新疆乌鲁木齐市谢家沟地区(试验所在地), 属于典型的山地草原, 哈萨克族牧民定居于此, 而混播草地是当地最主要草地类型之一, 传统的利用方式主要以割草(8月中旬)和放牧(9-10月)为主, 存在的问题:1)割草期过晚(一般在8月中旬), 大部分牧草已进入结实期, 牧草营养价值较低; 2)8和9月为干旱少雨期, 刈割以后牧草再生较困难, 导致9-10月人工草地无草可放牧。在这样粗放管理和过度利用下, 已建植的混播草地生产力极低, 稳定性较差, 难以保证家畜的冬季饲草料供给。而如何提高该地区混播草地的生产力和稳定性对于该地区畜牧业发展具有重要意义。针对以上问题, 本研究在调查了当地现有的混播草地面积、牧草产量、营养价值、利用方式的基础上, 选择了当地最常使用的4种主要混播牧草无芒雀麦、鸭茅、红豆草、红三叶的栽培种, 以单播为对照, 设置了2种豆禾组合和3种组合进行了混播, 并对建植混播草地的生产力及物种组成的变化长期监测。在本研究中, 混播草地于2016年6月播种, 当年形成的生物量不足以刈割晾晒干草, 仅在2016年底进行了放牧。2017和2018年6月底收获的牧草(刈割干物质产量)主要用于晾晒干草以备家畜冬天饲喂。从结果分析可知, 牧草单播, 2种牧草混播, 3种牧草混播的刈割干物质产量均表现出2017年显著高于2018年。如2017年4种牧草单播的平均干物质产量为2018年的1.83倍; 2017年2种牧草混播的平均干物质产量为2018年的1.39倍; 2017年3种牧草混播的平均干物质产量为2018年的1.51倍。这可能和牧草生长期间的有效降水量有关。2017和2018年4-6月降水量分别为147和111 mm(图1)。2016、2017和2018年混播草地刈割后再生的牧草干物质产量(第2茬)主要进行秋冬季(10月开始)绵羊和肉牛混合放牧利用。在放牧之前对其干物质产量、密度和粗蛋白产量进行测定。总体来看, 4种牧草单播, 2种牧草混播, 3种牧草混播的放牧平均干物质产量在年际之间均表现出2018年显著高于2016和2017年。这可能与2018年7月至9月降水量为171.4 mm, 较2016和2017年有所增加有关系。因为水分是影响草地生态系统生物多样性、生产力和稳定性最主要的因子[25, 26], 干旱可显著降低草原植物群落的物种丰富度和生物量[27]。
在混播草地中, 物种之间存在着对资源(养分、水分、空间和光照)的竞争, 竞争的结果将导致物种组成的变化。一般来讲, 竞争力强的牧草往往排挤竞争力差的牧草, 使得草地群落处于不稳定的状态[28]。一般认为, 氮和磷养分也是草地生态系统生产力和牧草品质的主要限制因子[29, 30]。在干旱半干旱地区, 水分和养分对草地生产力的影响是耦合的, 在干旱年份以水分限制为主, 而在多雨年份则转化为养分限制为主[26, 27, 28, 29, 30, 31]。在本研究中每年定期施磷钾肥, 可在一定程度上补充牧草生长过程中对土壤养分的消耗。该地区2016-2018年植物生长期间累计降水量分别为389.6、250.1和282.5 mm, 是典型的干旱半干旱地区, 水分可能是限制混播草地生产力的首要因素[32]。当土壤干旱时, 禾本科植物的须根系先感知缺水并抑制生长, 而豆科植物的直根系入土较深可吸收利用土壤深层水而满足生长需求, 根系越深在干旱环境中生长越有利。
在本研究中单播草地中无芒雀麦和红豆草干物质产量显著高于红三叶和鸭茅, 而在混播中, 有无芒雀麦和红豆草参与的混播草地干物质产量也显著高于其他混播组合。其原因主要是红豆草植株个体高, 茎秆粗壮冠幅大, 分枝密度较高, 这决定了其在空间和光照资源的竞争中占据优势[11]。相对其他3种牧草来说, 无芒雀麦是一种耐旱能力较强的根茎型牧草, 在同等条件下更能适应生境条件的变化[33], 其在混播草地和单播时具有较高的干物质产量可能与其有关。禾本科牧草叶片集中于下部, 豆科牧草叶片集中在中上部, 豆禾混播能充分利用地上空间和光资源提高生产力。因此, 不同的混播比例对混播草地的生产力也具有重要影响[33, 34, 35]。如陈积山等[36]分析了不同混播比例下紫花苜蓿(Medicago sativa)和无芒雀麦的牧草产量, 认为无芒雀麦20%+苜蓿80%混播组合牧草产量最高。当紫花苜蓿50%+无芒雀麦50%混播时, 混播草地产量介于二者单播牧草产量之间, 高于其平均值[37]。由此可见, 豆禾混播草地能显著提高牧草产量, 与禾本科牧草单播相比尤为明显。但是否高于豆科牧草单播产量受混播牧草种类、混播比例、生长年限、刈割茬次以及管理水平等多因素影响。
人工草地群落的稳定性是草地长期保持生产力的基础, 也是合理有效地建植、利用、管理及改良人工草地的基本依据[38]。混播中各物种干物质产量所占比例不仅反映各物种对混播草地生产力的贡献率, 还能间接反映出各物种对环境资源的竞争能力。混播草地中各物种比例的消长也可预测出混播草地未来的变化趋势, 间接地反映出混播草地群落的稳定性。无芒雀麦分别与红三叶和红豆草混播时, 无芒雀麦的干物质产量所占总干物质产量的比例随着年度的增加而显著增加, 相反, 红三叶和红豆草干物质产量所占比例显著下降, 特别是红三叶在建植后的第3年有从混播草地中消失的趋势。同样地, 鸭茅分别与红三叶和红豆草混播时, 鸭茅和红三叶干物质产量所占比例无明显的变化规律, 红豆草干物质产量所占比例显著增加, 而鸭茅的干物质产量却显著降低。这可以预测未来有无芒雀麦参与的混播草地中无芒雀麦干物质产量将占据绝对优势, 在与无芒雀麦的混播中红三叶将在建植的第4年从混播中消失, 而红豆草虽有下降的趋势, 但仍能在混播中占据一定的生态位。从3种牧草混播草地的干物质产量比例变化趋势中可以预测, 未来红豆草/红三叶/无芒雀麦和红豆草/无芒雀麦/鸭茅3种牧草混播草地将变成无芒雀麦/红豆草2种牧草混播草地, 红三叶和鸭茅将从该混播草地中消退, 这可能与红三叶和鸭茅相对较差有关系[39]。在本研究中分别与无芒雀麦和鸭茅混播时, 红豆草的竞争表现呈现出不一样的结果:随着年度的增加, 红豆草与无芒雀麦混播时其干物质产量所占比例逐渐下降, 无芒雀麦的比例逐渐增加, 在与鸭茅混播时, 红豆草的比例逐渐增加, 而鸭茅的比例逐渐下降。这说明在混播中红豆草产量的持久性主要受伴生牧草竞争能力的影响, 这与Malisch等[40]报道的结果一致, 该研究认为当建植混播草地在第3年时, 在与鸭茅混播草地中红豆草的比例占17%, 与黑麦草(Lolium perenne)混播草地中红豆草比例占38%。
株丛密度是反映草地生产力的一个重要指标。研究发现株丛密度与产量相关性极显著[41]。考虑到不同牧草其生物学特性, 在本研究中测定了牧草的分枝(分蘖)密度。随着生长年限的增加, 不论是单播还是混播中, 红三叶和鸭茅的密度均逐渐下降, 而无芒雀麦的密度逐渐增加。这说明从混播草地密度的变化来看, 随着年限增加, 无芒雀麦在混播草地中逐渐占据优势, 鸭茅和红三叶在混播草地中逐渐处于劣势, 且有消退的趋势。其主要原因可能与其生物学特性与环境适应性有关。这主要是因为在混播草地中物种之间竞争力的差异主要受物种形态差异的影响[42]。其中无芒雀麦作为一种根茎型多年生禾草, 具有根茎和种子两种繁殖方式, 一般在割草和放牧利用下, 其主要依靠无性繁殖来扩张。无芒雀麦的根茎型须根系可以保证该物种在浅层土壤中有效地吸收养分和水, 增加其与周围植物的竞争力[43]。陈积山等[36]研究也表明在无芒雀麦与苜蓿的混播中, 无芒雀麦的竞争力一直大于苜蓿。红豆草作为一种较抗旱的优良豆科牧草, 一般与抗旱性较强的禾草混播效果较好[44], 也因其含有浓缩单宁在放牧中不易引起家畜的鼓胀病[45]。与伴生的禾草相比, 在前期红豆草的竞争优势非常明显, 但是在后期竞争力也减弱, 容易被其他的竞争力强的物种取代[45]。Sanderson等[46]研究也认为红豆草不论是在单播还是在混播中其稳定性和持久性较差, 需要定期的重建以维护其稳定的生产力。可见红豆草更适合作为一种短期的轮作豆科牧草[47]。从无芒雀麦、鸭茅、红三叶和红豆草在单播和混播中生物量和密度的一致性来看, 生物量的变化与其密度的消长有密切关系。
混播草地牧草的营养价值主要取决于参与混播草地物种及其所占的比例, 混播草地的粗蛋白产量主要取决于各物种的干物质产量和植物氮含量。从本试验的结果来分析, 环境条件(特别是自然降水量)、物种适应性和物种对资源的竞争力通过影响混播草地各物种干物质产量和植物氮含量来影响混播草地牧草粗蛋白产量。随着年度的增加, 混播草地物种比例发生变化进而影响混播草地牧草的粗蛋白产量。
干旱区混播草地年际间干物质产量的消长变化主要取决于年均降水量和牧草的适应性。从混播草地干物质产量和密度的变化来看, 随着年限增加(3年), 无芒雀麦和红豆草在混播草地中逐渐占据优势, 鸭茅和红三叶在混播草地中逐渐处于劣势, 且有消退的趋势。关于该地区混播草地建植与管理的建议:1)从鸭茅和红三叶的表现来看当地建植混播草地时不建议选择鸭茅和红三叶; 2)该地区混播草地管理适合割草+放牧, 割草期应提前至6月底至7月初, 一方面保证了收获牧草的营养价值, 另一方面为秋冬季放牧牧草再生留够时间。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|