



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
荒漠草原土壤种子库对灌丛引入和降水梯度的响应特征
[于露1, **  , 周玉蓉
, 周玉蓉1, ** , 赵亚楠1 , 郭天斗1 , 孙忠超1 , 王红梅1, 2, * ]
, 周玉蓉, 赵亚楠]
|
|


作者简介:于露(1995-),女,吉林松原人,在读硕士。E-mail: yulu5890@163.com。周玉蓉(1994-),女,甘肃武威人,在读硕士。E-mail:1206525170@qq.com。
**共同第一作者.
为了揭示宁夏荒漠草原土壤种子库对灌丛引入过程及降水的响应特征,采用室内萌发法和野外水分激发法,对宁夏荒漠草原封育草地、放牧地、不同年限(3、12、22 yr)和间距(40、6、2 m)柠条灌丛地开展草地-放牧-灌丛引入过程的土壤种子库萌发及其对不同降水梯度(-60%,-40%,-20%,0,20%,40%)响应特征进行了研究。结果如下:不同年限、间距引入灌丛地0~200 cm年平均土壤水分含量显著低于封育草地和放牧地( P<0.05),随着灌丛年限和密度增加,加剧深层土壤水分的消耗,土壤旱化加重,使得灌丛地无明显春季返潮期,土壤水分仅为8.14%、8.83%,分别显著低于同期封育草地和放牧地16.00%、13.00%,在这样的水土条件下不同年限、间距灌丛地室内萌发下的0~10 cm土壤种子库密度分别为6186和5831 粒· m-2,分别高于封育草地、放牧地2965、2065粒·m-2( P<0.05)。各样地主要集中于0~5 cm土层,占萌发总数70.04%,且均以一年生猪毛蒿萌发为主,高达87.12%~96.55%,并随灌丛年限和密度的增大而增加,而多年生草本萌发种类仅为5种;不同降水梯度野外激发下的不同年限、间距灌丛地种子萌发平均数量为855和640粒·m-2,以降水量+20%时所激发土壤种子库萌发密度最高,分别为2266和1600粒·m-2,一年生与多年生草本种子库萌发密度和种类均随降水量升高而增加,且均在降水梯度最大时(+40%)呈下降趋势,而且相较于多年生草本,一年生草本在不同灌丛地萌发率高达90.58%~100.00%,显著高于放牧地或封育地( P<0.05),在灌丛引入过程中对土壤干旱和降水变化表现得更为敏感和适应。
**These authors contributed equally to this work.
This study explored the responses of the soil seed bank (0-5 and 5-10 cm soil depths sampled) to variation in precipitation and to encroachment by anthropogenically introduced shrubs ( Caragana) in desert steppe grassland in Yanchi County, Ningxia Province. Simulated rainfall levels relative to mean annual precipitation of 290 mm (-60%, -40%, -20%, 0, 20%, 40%) were generated for eight otherwise similar sites with different vegetation categories: grassland fenced to preclude grazing (FY), grazed grassland (FM), and grassland with introduced shrubs established for 3, 12, or 22 years (3 sites, PY3, PY12, PY22) or at planting distances of 2, 6, or 40 m (3 sites, PS2, PS6, PS40). Species represented in the seedbank and their numbers were determined using germination counts after incubation of sampled soil in the field and in the greenhouse. A major finding was that in grassland with introduced shrubs, soil moisture for 0-200 cm soil depth was 8.14% when averaged across seasons and over PY sites, and 8.83% when averaged over PS sites, which was significantly lower ( P<0.05) than corresponding values for FY and FM sites (16.00% and 13.00%, respectively). PS and PY sites also did not show higher soil moisture levels in spring than in autumn that were seen in FY and FM plots, and the soil moisture reduction at 40 m spacing was about 50% of that at 6 and 2 m shrub spacing. The number of species detected in the soil seed bank in field germination tests at the eight sites ranged between 4 and 7, comprising annual herbs, perennial herbs and shrubs, with total counts of 2965·m-2 and 2065·m-2, respectively, for FY and FM sites; 6198, 6465 and 5897·m-2, respectively, for PY3, PY12 and PY22 sites; and 3898, 5897 and 7698·m-2, respectively, for PS40, PS6 and PS2 sites. Of the total seed bank germination counts across the eight sites, 89.6% (range 87.12% to 96.55%) belonged to the annual herb Artemisia scoparia, and 70.04% came from the 0-5 cm soil layer. The number of species detected in glasshouse pot germination tests at different simulated rainfall levels, for samples from the eight sites, ranged between 1 and 5. For soil samples from all eight sites, the highest numbers of germinated seeds occurred in the +20% rainfall treatments and the lowest in the -60% treatment, while the +40% treatment was usually second lowest. For FY and FM sites the germinated seed density of the +20% treatment was 533 and 233·m-2, respectively. For sites with introduced shrubs, peak germination was 2266·m-2 and 1600·m-2 averaged over PY and PS sites, respectively. In summary, at these sites, introduced Caragana shrubs depleted soil moisture at depth, and the soil moisture reduction increased the dominance in the seedbank of the annual herb Artemisia scoparia, and tended to reduce species richness of the seedbank.
在过去的160年里, 木本植物的灌丛化已经影响了全球几亿hm2的干旱和半干旱草原地区。而引起荒漠灌丛化过程的一系列机制表明, 过度放牧、干旱、降水频率的改变、火烧、大气中CO2浓度的增加均可以解释该现象。已有学者通过对种子库及植被更新的预测研究得出连年干旱是美国西南部的迟花环地区灌丛迅速扩张的主要原因[1, 2, 3, 4]。土壤种子库(soil seed bank)作为荒漠草原上潜在的植物群落, 是种群生存、繁衍和扩散的基础, 是重建退化生态系统的重要组成部分[5], 但为加快退化荒漠草原恢复进程, 部分地区选择大规模引入灌丛作为恢复的重要措施[6], 这加速了草原土壤深层水分消耗, 加剧了草原干旱化程度, 影响了原生草本种子的萌发和定植, 减缓或抑制了原生草本的自然更新过程, 造成了草本植物多样性的减少, 对草原生态系统功能等方面造成了不可逆转的变化[7, 8]。
降水是影响干旱地区种子库组成和密度的最主要因素, 作为土壤有效水分的主要来源, 降水是荒漠草原草本植物发芽、生长以及生产的主要驱动力。随着全球气候变化背景下干旱地区出现降水频率低、暴雨频发等不规律降水模式的改变, 人为选择下的陆地上植被恢复率减少[9]。土壤种子库内一年生草本种子能够积极响应不同降水强度, 萌发比例占总种子库的70%以上[10], 而80%的干旱地区多年生草本植物为应对干旱带来的危机, 通过种子生殖并在土壤中休眠数年这一策略进行生存[11, 12]。草原灌丛化在时间和空间上都对土壤湿度有着一定的影响[13], 迟花环地区灌丛化的植被更新研究表明:相较豆科灌木下的土层, 草原在冬季明显吸收更充足的水分, 为春季植被萌发储存更多水分, 而灌丛下土壤水分蒸腾、消耗显著高于草地, 降低了土壤水分随降水的波动, 因此不同植被在长期干旱下土壤水分与土壤特性之间的变化会随季节而产生波动[14]。土壤干旱程度加深, 加之干旱、半干旱地区的有效降水频率逐渐减少, 将进一步减缓植物种类有效更新速度, 种子休眠现象难以缓解, 从而导致种子发育不良, 因此成为植被更新失败的主要原因[15, 16]。虽然干旱地区的多年生草本具有对抗长期干旱和水分匮缺的特性, 但是种子早期萌发、破土和幼苗生长易受土壤水分匮缺的影响, 进而影响整个地区的植被更新[17, 18]。目前, 国外相关学者通过研究种子休眠、萌发和定植对土壤水分的响应, 对灌丛化过程中土壤种子库以及草原植被更新机理等相关内容进行深入研究, 确定了多年生草种和入侵灌丛种子更新的年限以及更新所需的水土条件[13], 但在人为大面积引入灌丛进行植被恢复的荒漠草原内, 降水模式和灌丛引入对土壤湿度以及荒漠草原植被更新的影响这一领域仍需要进一步的研究和验证[19]。
基于此, 本研究开展不同年限和间距柠条(Caragana korshinskii)灌丛引入过程中宁夏荒漠草原的土壤种子库在不同降水梯度下的变化特征研究, 以期了解土壤种子库对不同灌丛引入过程以及降水模式变化的响应特征, 阐明灌丛引入对草本植被更新和植被演替动态影响, 为避免荒漠草原地区人工灌丛引入向大面积灌丛自我扩张发展以及荒漠草原地区植被重建与恢复可持续提供理论和数据支持。
研究区位于宁夏盐池县东北部花马池镇柳杨堡-十六堡一带(37° 05'-38° 10' N, 106° 30'-107° 39' E), 该地区属典型的温带大陆性气候, 海拔1380~1600 m, 由于受到来自蒙古-西伯利亚的高压、冷湿空气的影响, 气候具有夏炎冬寒、降水稀少、全年降水分布不均匀、蒸发量大等的特征, 年平均气温8.1 ℃, 无霜期160 d, 年平均降水量250~350 mm, 降水集中在7-9月, 年平均蒸发量2139 mm, 平均年风速为2.8 m· s-1。该地区植被类型为荒漠草原、沙生植被及大量的人工灌丛, 草本植物主要有:猪毛蒿(Artemisia scoparia)、白草(Pennisetum centrasiaticum)、草木樨状黄芪(Astragalus melilotoides)、糙隐子草(Cleistogenes squarrosa)、狗尾草(Setaria viridis)、米口袋(Gueldenstaedtia verna)、乳浆大戟(Euphorbia esula)、砂珍棘豆(Oxytropis racemosa)、雾冰藜(Bassia dasyphylla)、画眉草(Eragrostis pilosa)、远志(Polygala tenuifolia), 灌木、半灌木主要是锦鸡儿(Caragana korshinskii)和牛枝子(Lespedeza potaninii)。土壤类型以灰钙土为主。
在盐池县东北部(十六堡-柳杨堡)分别选取封育草地(FY)、放牧地 (FM, 对照)、不同种植年限(3、12、22 yr)柠条灌丛、不同间距(40、6、2 m)柠条灌丛(均为22 yr), 利用空间代替时间的方法来代表荒漠草原退化及人工灌丛引入恢复的不同阶段和过程, 即:1)草地-放牧-不同种植年限灌丛引入过程为FY、FM、PY3(种植3年)、PY12(种植12年)及PY22(种植22年)(PS6); 2)草地-放牧-不同种植间带灌丛引入过程为FY、FM、PS40(种植间距40 m)、PS6(种植间距6 m)和PS2(种植间距2 m)(图1, 表1)。
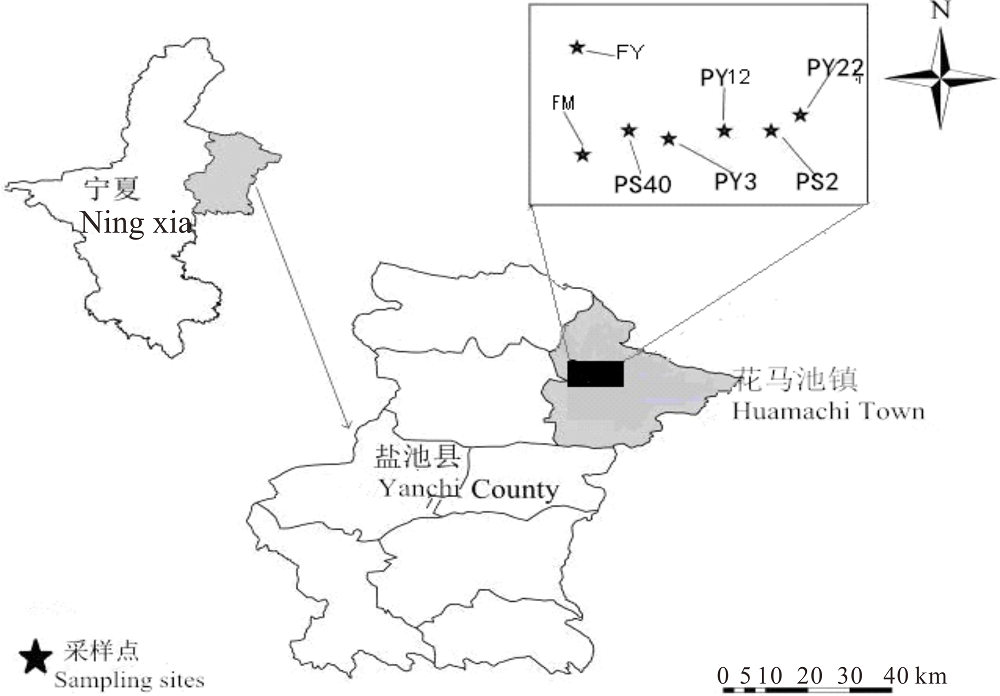 | 图1 样地采样点分布Fig.1 Sample points of sampling plots |
| 表1 样地概况 Table 1 Sample overview |
1.3.1 土壤水分测定 分别在封育草地、放牧地随机设置5个重复观测点, 不同种植年限、间距柠条灌丛地在带间、带内分别设置3个重复, 进行定点观测, 采用时域反射仪(time domain reflectometry, TDR)对土壤水分进行观测, 观测深度为0~200 cm, 每20 cm为一层, 共10层, 观测时间为2016、2017年3-11月, 每月1~2次。
1.3.2 土壤种子库取样与萌发 2016年11月、2017年3月在确定的样地进行土壤种子库随机取样, 采样时避开样地边缘30 m, 设置封育地、放牧地、柠条灌丛样地(分别在带内、带间)各选取5个面积为1 m× 1 m的采样方, 每个样方内设置2个采样点, 采用10 cm× 10 cm× 10 cm取样器, 对每个采样点分别进行0~5 cm、5~10 cm两层取土。室内萌发试验则是将取回的土样先除去大的砾石和粗根系, 之后将土样均匀铺设在内径14 cm、高15 cm塑料花盆(塑料花盆底部预先填上5 cm厚的无种子的蛭石)中。因考虑到有些植物在3 cm以下的土层中难以破土萌发, 因此铺设在蛭石上的土样厚度应控制在3 cm以内, 最后在适宜温度环境(温度20~25 ℃)下进行萌发试验。根据前期研究土壤水分得出研究区田间持水量为21.26%± 1.89%, 因此将22%作为基准含水量进行适时浇水, 时间在6周左右, 添加40 mg· L-1营养液· 月-1(5 mL营养液/10 L水, N∶ P∶ K=1∶ 1∶ 1), 同时记录每周各处理的发芽情况以及后期的植物鉴定工作, 待幼苗生长到形态特征比较明显时进行植物统计和种类鉴定, 秋、春季土壤种子库萌发一般持续到3和7月(该试验中春季种子经过冬季低温过程, 萌发数量种类均高于秋季采样, 结果中的统计均以2017年3月采样观测分析为主)。
1.3.3 土壤种子库降水野外激发试验 根据研究区多年平均降水量290 mm为基准, 以确定不同年降水量梯度对土壤种子库的激发效应, 从而研究不同降水梯度对土壤种子库物种组成和总萌发数量的影响, 降水梯度设置为多年平均降水量的-60%、-40%、-20%、0、20%、40%, 即降水量分别为116、174、232、290、348、406 mm, 分别对以上7个样地(PY22和PS6为同一样地)盆栽中土壤水分进行定期观测并添加定量水分(由降水量梯度总含量、计划施水次数与盆栽器皿直径决定)以保持土壤湿度, 每个样地均有3个重复。添加水分的周期为5 d, 施水量分别为74、111、148、185、223、260 mL· 次-1, 为避免自然降水对试验造成的误差, 进行遮雨措施。该试验于2017年3月开始, 每周记录各样地盆栽内的发芽情况, 当幼苗生长到具备明显的形态特征时, 进行植物统计和物种鉴定。萌发时间持续4个月, 大约为120 d。若20 d内无新幼苗萌发, 则认为萌发结束。
依据采样时所取土的面积, 将盆栽中记录萌发出的幼苗数换算成每m2的幼苗数, 以此来表示土壤种子库的大小, 即为土壤种子库密度[5]。
采用SPSS软件中的单因素方差分析(One-way ANOVA)对不同样地土壤种子库物种萌发总密度进行统计分析, 用最小显著差异法(LSD)进行多重比较检验, 显著水平为P< 0.05。同时采用Excel 2010、Origin Pro 8.0软件进行作图。
在草地-放牧-不同年限灌丛引入过程中(图2), 各灌丛样地土壤水分含量范围为6.99%~11.61%, 显著低于FY和FM的平均含量14.91%、13.14%(P< 0.05); FY、FM土壤水分季节动态表现为春季返潮上升期到夏季生长消耗降低期(7-8月)进而秋季雨水补充增加期的变化趋势, 而不同年限引入灌丛地表现为春季返潮时土壤水分深层效果较差, 平均土壤水分仅为8.14%, 显著低于同期封育草地、放牧地16.00%、13.00%; 各个样地土壤水分均在7-8月呈低谷值, 为7.70%~9.31%; 随着秋季降水的增加各样地土壤水分均呈上升趋势, 但不同年限引入的灌丛草地土壤水分仍显著低于FY和FM(P< 0.05)。
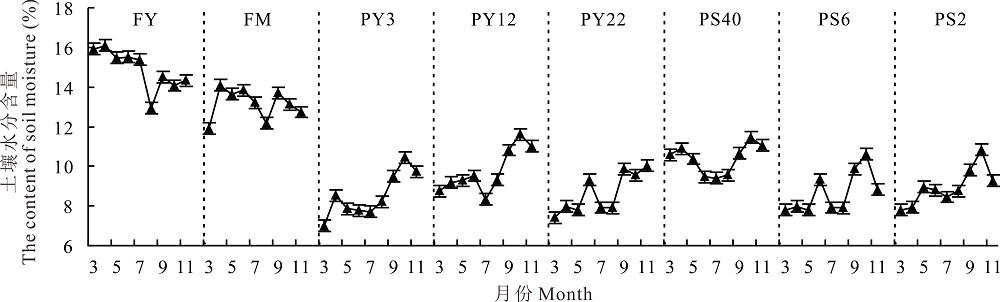 | 图2 不同年限和间距灌丛引入后土壤水分含量季节动态变化Fig.2 Seasonal dynamic of soil water content in desert grasslands after shrub colonization in different ages and spaces |
草地-放牧-不同间距灌丛引入过程中(图2), 各灌丛样地土壤水分含量范围处于7.80%~11.45%, 显著低于FY和FM (P< 0.05); 由于深层次土壤水分消耗较多, 使春季返潮期内不同间距灌丛地土壤水分较低, 为7.80%~10.90%; 各个样地土壤含水量均在7-8月处于最低值, 为7.90%~9.56%。随着灌丛引入密度的增加, 土壤含水量呈降低趋势, 显著低于FY和FM(P< 0.05)。
2.2.1 土壤种子库密度及物种组成特征 随着不同年限灌丛引入过程(表2), PY12土壤种子库萌发密度最高, 为6465粒· m-2; 各样地中菊科(Compositae)的猪毛蒿密度均为最高; PY22、PY12和PY3与FM和FY之间存在显著差异(P< 0.05); FM多年生草本所萌发密度最高, 达399粒· m-2。各样地间一年生草本萌发密度显著高于多年生草本, 且PY3、PY12及PY22内一年生草本数量明显高于FY、FM。
| 表2 不同年限和间距引入灌丛地土壤种子库物种组成及密度 Table 2 Species composition and density of soil seed bank for different ages and spaces (No.· m-2) |
在不同间距引入灌丛过程中(表2), PS2土壤种子库密度最高, 为7698粒· m-2; 各样地中猪毛蒿密度均为最高; PS2、PS6和PS40均与FM、FY之间存在显著差异(P< 0.05); 其中, FM内多年生草本土壤种子库密度最高, 为399粒· m-2。
2.2.2 土壤种子库垂直分布 随着不同年限灌丛引入过程(图3), PY22、PY12和PY3与FM、FY土壤种子库密度在两个土层中总体差异显著(P< 0.05); 在不同间距灌丛引入过程下, PS2、PS6和PS40与FM、FY土壤种子库的密度分别在两个土层中总体差异显著(P< 0.05)。在两个引入过程中均以0~5 cm土层种子库萌发密度最高, 约占总萌发密度的70.04%。
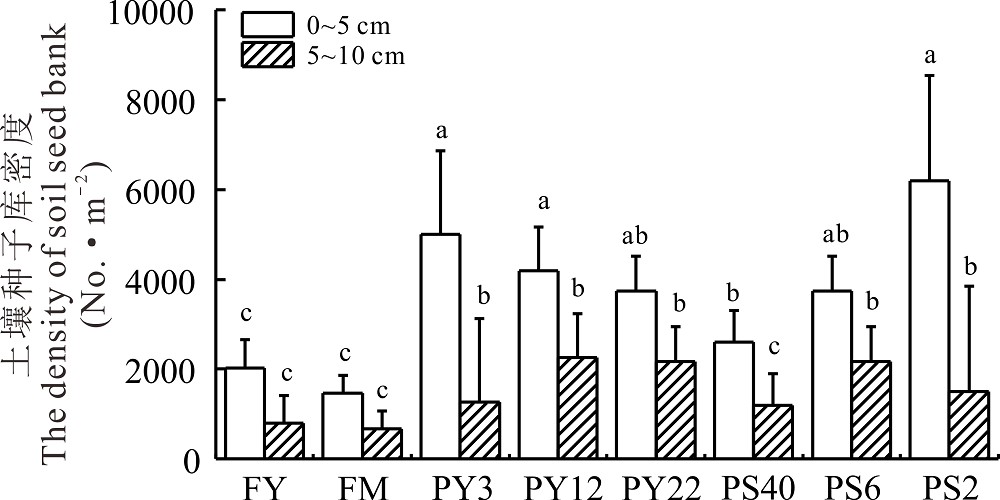 | 图3 不同年限、间距引入灌丛地土壤种子库垂直密度分布 不同字母代表不同处理间差异显著(P< 0.05)。 下同。Fig.3 Vertical distribution of seed bank densities in introduced shrublands with different ages and spaces Different lowercase letters indicate significant differences between the different treatment (P< 0.05). The same below. |
2.3.1 不同灌丛引入过程中萌发种子库对降水梯度的响应 随着降水量的递增(图4), 土壤种子库密度变化表现为先增加后逐渐减少的趋势。随着降水量的增加FY与FM较PY3、PY12、PY22种子库密度变化较缓慢; 降水量在348 mm时, 各样地土壤种子库中可萌发种子密度最大, 且差异显著(P< 0.05), 其中一年生草本猪毛蒿为优势种, 占总数的92.71%; 当降水量为116 mm时, 种子库萌发密度最低。6个不同降水水平下的不同年限引入灌丛地内土壤种子库物种数变化相对平缓, 随降水量的递增物种数量呈先增加后降低的趋势, 且均以猪毛蒿为主。
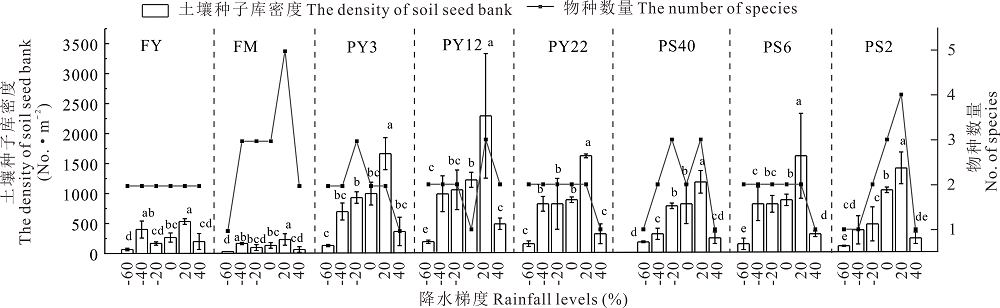 | 图4 不同降水梯度对不同年限和间距引入灌丛地土壤种子库萌发密度和数量激发Fig.4 Densities and size of seedlings from soil seed banks in different ages and spaces of introduced shrublands fields at different rainfall levels |
不同间距引入灌丛地内土壤种子库密度变化情况表现为先增加后减少的趋势(图4), FY与FM较PS40、PS6、PS2种子库密度变化较缓慢; 降水量为116 mm时, 土壤种子库中可萌发种子数最低; 年降水量在348 mm时则激发密度最高; 且仍以优势种猪毛蒿为主, 占种子萌发数量的92.80%。
2.3.2 不同生活型植物种子库对降水梯度的响应 不同年限、间距下引入灌丛地一年生草本种子库萌发密度随降水量(116~348 mm)升高呈显著增加趋势(P< 0.05), 多年生草本种子库萌发密度随降水量也呈增加趋势(P< 0.05), 但在降水量最大时(406 mm)均出现显著下降变化(图5)。在降水量为348 mm时两者均达最大值, 一年生草本密度分别为2100、1600粒· m-2, 多年生草本密度各自为166、100粒· m-2; 当降水继续增大到406 mm时, 各灌丛地一年生和多年生草本萌发密度均呈显著下降, 此时一年生草本密度范围为0~466粒· m-2, 而多年生草本萌发密度范围为0~33粒· m-2。灌丛引入过程中种子库萌发对降水的响应仍以一年生草本为主(表3), 其萌发率各自占总数的95.42%、97.37%。
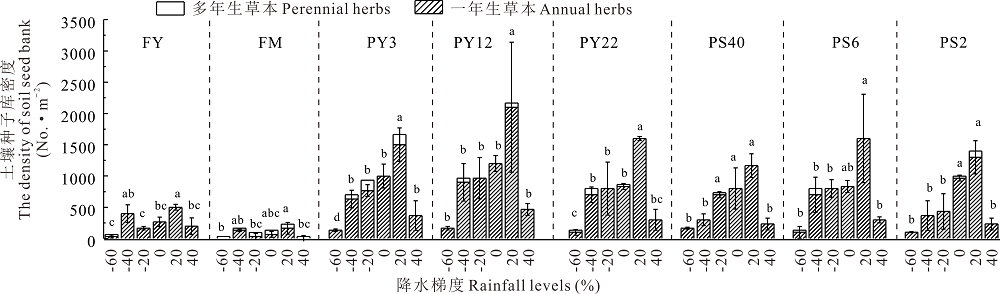 | 图5 不同年限和间距引入灌丛地不同生活型植物种子库对降水梯度的响应Fig.5 Responses of soil seed banks of life forms to different rainfall levels in introduced shrubland fields with different ages and spaces |
| 表3 不同生活型植物对不同降水梯度回归模型 Table 3 Correlation analysis of different life forms under different rainfall levels |
土壤水分作为干旱、半干旱地区最为主要的限制性因素, 在夏季变化剧烈、在冬季则变化缓慢[15], 并有研究指出与总降水量相比, 季节性降水对生产力影响更大[20]。正如本研究中封育草地、放牧地0~2 m土壤水分结果所示, 其季节动态与春季返潮期-夏季消耗期-秋季补充期和植物生长、降水动态相对应, 虽有滞后, 但基本相似。这是由于夏季植被生长需水较强, 除地表蒸发外, 还有地上植被自身所需以及植物蒸腾作用, 加快了土壤水分消耗, 而冬季植被生长缓慢、地表蒸发量减少。但在不同年限、间距引入的灌丛地内, 明显存在春季土壤含水量显著低于封育草地和放牧地的现象, 这是由于春季土壤返潮水分主要来自前一年秋季的降水和深层土壤水分的充足补给, 但因灌丛具有强大的主根系和发达的侧根系, 在生长中会更多利用深层土壤水分, 以及浅层处0~30 cm处水分[21], 因此造成土壤水分匮缺严重, 春季土壤返潮效果这一现象不明显, 进而导致春季土壤水分不足。
因土壤种子库与生存环境有密切联系, 土壤水分作为土壤特性的重要指标对土壤种子库萌发有着重要作用[22, 23]。在时间和空间尺度上, 灌丛化会对土壤表层水分、能量、营养物质以及碳循环等有影响, 也因其较强的持水及贮存土壤有机碳能力, 从而导致“ 沃岛” 效应[24]。与周围环境相比, 沃岛之外的草地土壤水肥较低, 即使春季返潮期补给一定的水分, 但因灌丛所导致的土壤水分空间异质性[25], 使得无法适应当前土、水条件的物种枯萎死亡, 从而降低土壤种子库密度及丰富度, 进一步影响种子萌发。本研究中种子库所萌发物种以一年生草本猪毛蒿为主, 且大多分布在0~5 cm土层, 而多年生草本种子库大多分布在5~10 cm, 这一结果则与李秋艳等[22]的研究结果相符合。随着灌丛引入年限和间距的增加, 土壤种子库萌发密度呈增加趋势, 即使在引入22 yr和2 m间距(密度最大)灌丛地出现下降, 但仍高于封育草地和放牧地, 这也与前人研究中随着不断演替, 早期物种及丰富度逐渐减少, 但种子库密度却未见减少[26]相符合, 这可能是一年生物种对荒漠区域中生物和非生物环境因素适应性较强, 更容易从干扰中恢复并进行再次繁殖[27]。同时相对宽阔的草原更容易受到因环境变化所引发的干旱影响[28, 29], 而相对密集的灌丛会给予种子以充足营养和保护, 充当“ 避风港” 拦截种子, 同时保护种子不被采食。封育草地种子库在萌发总密度和物种数上均处于低值, 一年生草本萌发密度也低于其灌丛地, 除了上述原因外, 也可能是在14年的围封下干扰降低, 地上群落仍以无性繁殖为主, 地上生物多样性较高, 物种的种子更新受干旱或休眠等因素影响变得缓慢或萌发不显著, 从而影响封育草地植被种子更新的周期[13]。
全球环境变化所引发降水格局的改变, 造成大部分区域会发生“ 干旱愈加干旱” “ 湿润愈加湿润” 现象[30], 通过模拟降水量逐渐增多(116~348 mm), 发现所激发的土壤种子库物种总数呈先上升后下降(406 mm)的趋势, 这与赵晓英等[31]的结果相符合; 在模拟年降水量最低为116 mm时可激发猪毛蒿种子萌发这一结果与闫建成等[32]的研究相近, 同时一年生草本密度均高于多年生草本, 且以一年生草本土壤种子库密度上升趋势最为显著, 多年生草本种子库萌发虽也呈增长趋势, 但仍低于总体萌发密度, 这说明降水增加可改变地上物种组成[33], 也能够促进更多物种能够存活, 以增加种子密度及丰富度。而干旱地区的物种为应对其生存环境, 主要有两种适应对策:一为干旱逃避, 多体现在一年生物种大量种子的繁殖; 二为干旱耐受, 指多年生物种以地下芽萌发、休眠等特性来规避干旱[34]。而相较于多年生物种来说, 一年生植物具有较好的适应对策, 如休眠及花期等生理特性使其能够在资源匮乏的环境中仍能完成其生活史[20], 同时, 对降水变化极为敏感且迅速, 能够有效利用水资源来调节自身的生长周期, 从而使之更容易应对降水多变的干旱环境。对于营养和水分贫乏的环境, 植物结构变化更倾向于短命和多变的趋势, 所以荒漠草原地区的一年生植物能够积极应对气候变化, 而在多变的降水条件下多年生植物种子更新难以准确预测[35]。在本研究中多年生草本种子可能由于干旱产生的应对机制发挥作用, 或是长期灌丛引入和气候变化下的土壤干旱使得多年生种子生活力下降(或已失去生活力), 不仅限制草原内潜在多年生种子库恢复, 同时也影响种子休眠和萌发[36], 而萌发过程还受自身遗传特性[如本研究中多年生草本扁蓄豆(Melissitus ruthenicus)种皮厚而硬]、生物结皮、温度以及动物采食等影响[37, 38, 39]。
由于荒漠草原人工引入灌丛过程加速了深层次土壤水分消耗, 使在种子萌发重要时期的春季土壤处于极端干旱状态, 而多年生草本又由于多种原因保持休眠或水分达不到萌发需要, 直接导致了多年生草本干旱耐受或失活退化, 一年生草本进一步积极响应。在全球降水模式逐渐向少量、长期干旱发展下, 本研究中的人工灌丛化及植被更新过程(一年生和多年生草本的萌发特征)直接响应上述变化趋势, 即灌丛引入的土壤旱化和气候干旱双重因素加剧了荒漠草原草本的退化, 使得干旱地区越加干旱, 植物群落变化向着一年生草本为主发展[40, 41], 导致荒漠草原区域多年生草本种子库萌发失败, 并进一步促进灌丛化过程中的一年生草本为主的植物群落演替。
荒漠草原灌丛引入过程中表现为灌丛地土壤水分均显著低于封育草地、放牧地, 虽各样地土壤水分均经历春季返潮期-生长消耗期-秋季降水补充期的季节动态, 但因灌丛引入加速了土壤深层的水分消耗, 使得不同年限和不同间距引入的灌丛地3-4月春季返潮效果较弱, 对春季种子萌发产生影响。在灌丛引入过程中一年生草本较多年生草本对干旱和降水变化更为敏感和适应, 在全球气候变化下的荒漠草原人工灌丛引入加速了植被演替向着短命、多变植物群落方向发展。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|