

{kind=link}
{kind=link}
{kind=link}
甘肃内生真菌基因组密码子使用的偏好性分析
[李秀璋1, 2  , 宋辉
, 宋辉3 , 张宗豪1 , 徐海峰1 , 刘欣1 , 李玉玲1 , 李春杰2, * ]
, 宋辉]
|
|


作者简介:李秀璋(1987-),男,甘肃庆阳人,助理研究员,博士。E-mail: xiuzhang11@163.com
在翻译过程中,三联体密码子指导氨基酸肽链的合成。长期进化过程中,有些氨基酸对应多个三联体密码子,形成了密码子兼并性。不同物种的基因组中,一些氨基酸偏好使用某种密码子,形成密码子使用的偏好性。突变和自然选择压力是影响密码子使用偏好性的两个主要因素。甘肃内生真菌是一种重要的禾草内生真菌,其主要侵染宿主醉马草,一方面能提高宿主醉马草对生物与非生物的抗性;另一方面产生的有毒生物碱类次生代谢产物毒害家畜。以甘肃内生真菌核基因组为研究对象,通过分析该基因组的基本组成及密码子使用模式,确定最优密码子。同时对各密码子相关指标与基因长度、氨基酸的使用频率等进行相关性分析,明确影响基因组对密码子使用偏好性的主要因素。结果显示,GC3的分布范围主要集中在40%~60%,62.7%的基因有效密码子数使用频率分布在0~0.1,95.4%的有效密码子数低于期望值而且位于期望曲线的右下方,这意味着突变压力是影响甘肃内生真菌密码子使用偏好性的因素之一。然而,中性分析结果显示,GC12和GC3之间呈显著正相关( r=0.110, P<0.01)、回归线斜率为0.2729,表明突变对密码子使用偏好性的影响仅为27.29%,其他因素的影响所占比例约为82.71%。相关性分析结果显示,疏水性氨基酸对甘肃内生真菌密码子使用偏好性的影响系数最大,其次是芳香族氨基酸。此外,在甘肃内生真菌基因组中总共鉴定到26个以C或G结尾的最优密码子,表明甘肃内生真菌偏好使用的密码子可能与第3位的GC含量有关。
The sequence of multiple sets of triplet adjacent DNA or RNA nucleotides (codon) determines amino acid sequence. In the long process of adaptive evolution, the codon usage of organisms has developed certain biases. Two major factors affect codon usage bias. The species ‘ Epichloё gansuensis’ is an important endophytic fungus. E. gansuensis infection can improve the host resistance to biotic and abiotic stress, but the fungus produces secondary alkaloid metabolites that may poison livestock. This study aimed to analyze the basic composition of the genome of E. gansuensis and the pattern of codon usage, and to determine the optimal codon. Correlation of each codon index with gene length, amino acid usage frequency and other indicators was analyzed, to identify the main factors affecting codon usage. We found that the GC3 is mainly distributed between 40%-60%, the effective codon number of 62.7% genes distributed between 0-0.1, 95.4% effective codon numbers are lower than the expected and lie to the lower right of the expectation curve. These results suggest that mutation pressure is one of the main influencing factors on codon usage of E. gansuensis. However, the neutral analysis showed a significant positive correlation between GC12 and GC3 ( r=0.110, P<0.01), with a slope of the regression line 0.2729. It suggests that the effect of mutation on codon usage was only 27.29%, 82.71% for other factors. Correlation analysis also showed that hydrophobic amino acids had the greatest effect on the codon usage of E. gansuensis, greater than that of aromatic amino acids. In addition, 26 optimal codon that end with a G or C residue from E. gansuensis were identified, indicating that codon usage in E. gansuensis may relate to the presence of a GC residue at the third position of the triplet codon. These optimal codon are significantly correlated with transcription level, and may help in the design of degenerative primers, for tracing the evolutionary origin of endophytic fungi.
真核生物的20种氨基酸对应61个三联体密码子。因此, 某些氨基酸由一个或多个密码子编码合成。其中, 编码合成同一种氨基酸的不同密码子被称之为同义密码子。同义密码子并不随机的同时出现在同一个基因组中, 因此, 这种现象被称之为密码子使用的偏好性选择[1, 2]。密码子使用偏好性的不同可以调节差异蛋白质生产的高效性和准确性, 同时保证同一条蛋白质序列的一致性。研究证实, 包括核苷酸组成的偏好性、突变、自然选择或选择翻译的优化都有可能影响密码子的使用模式。因此, 对某一物种的密码子使用模式进行分析, 可以解释其基因调控的分子生物学原理、基因表达、蛋白质二级结构、选择性转录和外部环境因素之间的相关性等[3, 4, 5, 6]。目前对于遗传密码子偏好性的研究主要集中在一些模式物种, 包括酿酒酵母(Saccharomyces cerevisiae)、登革热病毒(dengue virus)、大肠杆菌(Escherichia coli)、黑腹果蝇(Drosophila melanogaster)、拟南芥(Arabidopsis thaliana)和人类等[1, 7, 8, 9, 10, 11, 12, 13, 14]方面, 然而对真菌的研究相对较少。
禾草内生真菌是一类内寄生的专性寄生真菌, 常与禾本科(Poaceae)早熟禾亚科(Pooideae)的冷季型禾草形成互惠互利的禾草-内生真菌共生体[15, 16]。这种互惠共生系统通过产生的一些生物碱类的次生代谢产物可以给宿主禾草提供一系列的生物保护作用, 提高宿主禾草新陈代谢[17], 同时其产生的部分生物碱类物质是导致家畜中毒的主要原因[18]。甘肃内生真菌(Epichloё gansuensis)是研究禾草内生真菌的一种重要模式材料, 该菌于2004年首次被Li等[19]分离鉴定并命名[16]。甘肃内生真菌常与芨芨草属(Achnatherum)植物醉马草(Achnatherum inebrians)形成互惠共生的醉马草-内生真菌共生体, 内生真菌的侵染能提高宿主醉马草对多种生物与非生物胁迫的抗逆性[20, 21, 22, 23, 24]。目前相关研究主要集中在生物与非生物胁迫条件下共生体的响应及其机制, 然而这些研究大多关注于其生理生化水平, 相关互作的分子机制研究仍相对缺乏。
目前, 甘肃内生真菌菌株E7080的基因组已经完成测序并释放[25], 这为研究该类内生真菌分子生物学提供了较大的便利性。本试验拟以甘肃内生真菌基因组为研究对象, 分析密码子的使用模式及其偏好性, 并对导致密码子产生偏好性的相关因素进行分析, 以期对禾草内生真菌分子生物学及其与宿主植物互作的分子机制研究提供借鉴。
甘肃内生真菌(E7080, version 2)的全基因组测定由美国肯塔基大学真菌基因组工程完成(http://csbio-l.csr.uky.edu/endophyte/), 基因组编码序列(coding sequence, CDS)下载于GenBank数据库(http://www.ncbi.nlm.nih.gov)。为了提高供试基因组序列的质量并将抽样错误降到最低, 对于CDS序列的过滤主要基于以下几条原则:1)每条CDS序列的开始必须存在起始密码子并在结尾必须出现终止密码子; 2)每条CDS序列的长度不得低于300 bp; 3)检测到的重复序列(必须为精确匹配)必须被排除在有效的数据集之外。经过筛选之后, 最终共有7969条CDS序列用于密码子使用模式分析。
GC含量指的是每条CDS序列中碱基为G或C的总含量比例, GC1、GC2和GC3分别指的是密码子第1、2和3位碱基为G或C的含量比例。GC3s是指GC核苷酸在密码子第3位编码同一种氨基酸的同义密码子, 是评估密码子使用偏好性的一个重要参数。同样的A3s、G3s、C3s、T3s和GC3s指的是密码子第3位分别以这些碱基结尾的同义密码子使用的偏好性。GC12指的是密码子第1、2位为碱基G或C所占比例的平均值, 该指标用来进行中性分析。
密码子适应指数(codon adaptation index, CAI)常通过已知的高表达基因作为参照, 用来分析待测基因或基因组的密码子使用偏好性[26]。它的取值范围在0~1, 取值越接近1则说明基因的表达水平越高, 反之亦然。
有效密码子数(effective number of codon, ENC), 其取值范围通常在20~61, 常用来进行单个基因密码子偏好性的分析。这个指标同样也是用来测定一个蛋白质中所有氨基酸不均匀使用的偏差程度。ENC值常受到基因组组成的影响, 对于每个氨基酸, 如果编码该氨基酸的所有密码子被均匀使用(完全随机使用), 则ENC值为61, 但是对于每个氨基酸总定向使用同样一个密码子, 那么它的ENC值则为20[27]。相对同义密码子使用频率(relative synonymous codon usage, RSCU), 是对于一套同义密码子在观测到的使用频率与理论使用频率之间的比值。RSCU的取值平均等于1, 表明密码子的使用未发生偏差; RSCU值小于1, 表明观测到的值小于期望值, 反之亦然[28]。
亲水性平均值(general average hydropathicity, Gravy)指在一个基因产物中所有亲水性氨基酸的总和除以序列中的残留[29]。Gravy值越偏向于负值, 则说明蛋白质的亲水性越强; Gravy值越偏向于正值, 则说明蛋白质的疏水性越强。芳香值(aromo values, Aromo)指假设基因翻译结果中芳香族氨基酸[苯丙氨酸 (Phe)、酪氨酸 (Tyr)、色氨酸 (Trp)]所占的比例。Aromo值和Gravy值用来量化氨基酸的组成中主要的相关性趋势[30]。
ENC分析(ENC vs. GC3s)是分析同义密码子使用偏好性的重要参数, 可以直观地展示基因密码子偏好性使用的主要特征。ENC值的取值范围为20(对于每一个氨基酸只有一个密码子被有效使用)至61(密码子被均匀使用)。期望ENC值参照下列的公式进行计算[27]:
ENCexp=2+
式中:S指的是碱基GC含量(例如, GC3s)。
本研究所有基因的有效密码子使用频率及其期望值, 通过公式(ENCexp-ENCobs)/ENCexp绘制分布频率图, 评估有效密码子数的观测值与期望值之间的相关性, 其中ENCexp指的是有效密码子的期望值, 而ENCobs指的是有效密码子数的实际观测值。
中性分析[31]用来分析影响密码子使用模式及偏好性的主要因素, 主要包括评估和描述GC12和GC3之间的相关性。当中性分析的斜率为0时, 表明没有受到定向突变压力(完全选择)的影响; 当斜率为1时, 表明受到中性选择[31]。
最优密码子使用指数可用来代表基因表达水平。本研究通过ENC值的大小对所有CDS序列进行排序。分别从已排序数据集的前端和后端各取5%数据作为“ 高表达” 和“ 低表达” 两个数据集:其中“ 高表达” 数据集指的是整体ENC值中相对表达较高的基因, 表示其观测到的实际ENC值远离期望值, “ 低表达” 数据集则指的是整体ENC值中相对表达比较低的基因[32]。两个数据集中每条CDS序列中对应的“ 高表达” 和“ 低表达” 的RSCU值之间的差异(Δ RSCU)如果大于0.08, 则其对应的密码子被定义为最优密码子[33]。
相关性分析(correspondence analysis, COA)是广泛运用于密码子使用模式的多元统计分析的方法。虽然总共有59个同义密码子(除去编码甲硫氨酸Met和色氨酸Trp密码子), 为了生成一致的基于RSCU的相关性分析, 将其自由度降低至40之后消除差异归因于氨基酸的不平均使用[34]。
主要使用的软件有Mobyle server的在线分析(http://mobyle.pasteur.fr), 密码子一般指标分析软件Codon W (Ver.1.4.4) (http://codonw.sourceforge. net/culong.html# CodonW), 本试验中选择作为分析模型进行比对分析。CHIPS(http://mobyle.pasteur.fr/cgibin/portal.py#forms:chips)和CUSP(http://mobyle.pasteur.fr/cgi-bin/portal.py# forms:cusp)软件被用来计算密码子使用偏好性的指标。
供试基因组密码子使用的模式与其他指标之间的相关性使用SPSS 19.0 (SPSS Inc., Chicago, IL, USA)进行分析。使用Tukey-Kramer检验对相关性分析中的多重比对分析进行校正, 分别使用置信区间为P< 0.01和P< 0.05进行显著性分析[35]。其他指标的方差分析均使用GraphPad Prism 5 (GraphPad Software, San Diego, CA, USA)软件和SPSS 19.0 (SPSS Inc., Chicago, IL, USA)软件执行。
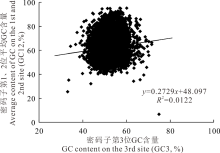
经筛选, 在甘肃内生真菌基因组中共得到7969条CDS序列, 平均GC含量为55.18%, 各序列的GC含量在30.60% 至73.10%之间(表1)。密码子3个位置上的平均GC含量分别为:57.84%(GC1)、45.25%(GC2)和62.16%(GC3), GC12的分布范围为40.00%~80.00%(图1)。
| 表1 甘肃内生真菌E. gansuensis密码子的基本组成信息、ENC、Gravy和Aromo Table 1 Base composition, ENC, Gravy and Aromo of codons of E. gansuensis |
 | 图1 甘肃内生真菌E. gansuensis中性分析(GC12 vs. GC3) GC12指的是密码子第1位和第2位的GC含量(GC1和GC2)的平均值; 而GC3代表的是密码子第3位上的GC含量; 实线代表的是GC12和GC3之间的线性回归, R2=0.0122, P< 0.01。Fig.1 Neutrality plots (GC12 vs. GC3) of E. gansuensis GC12 stands for the average value of GC content in the first and second position of the codons (GC1 and GC2). While GC3 refers to the GC content in the third position. The solid line is the linear regression of GC12 against GC3, R2=0.0122, P< 0.01. |
中性分析主要揭示GC12和GC3之间的相互关系(图1), 能够反映甘肃内生真菌基因组密码子使用模式中自然选择和突变压力之间的均衡。结果表明, 该基因组中GC3含量的分布范围在40%~60%, 密码子中GC12和GC3之间呈极显著正相关(r=0.110, P< 0.01), 回归线斜率为0.2729。
甘肃内生真菌基因组的有效密码子数(ENC)的范围为26.97~61.00, 其平均值为52.56。在筛选出的所有7969条基因中, 仅有166条基因表现出较高的密码子偏好性(ENC< 35), 表明该基因组对密码子使用的偏好性较弱。
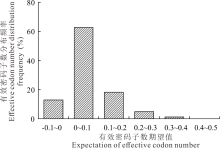
大多数基因有效密码子数的使用频率分布在0至0.1之间, 表明大部分有效密码子数的观测值小于期望值(图2)。
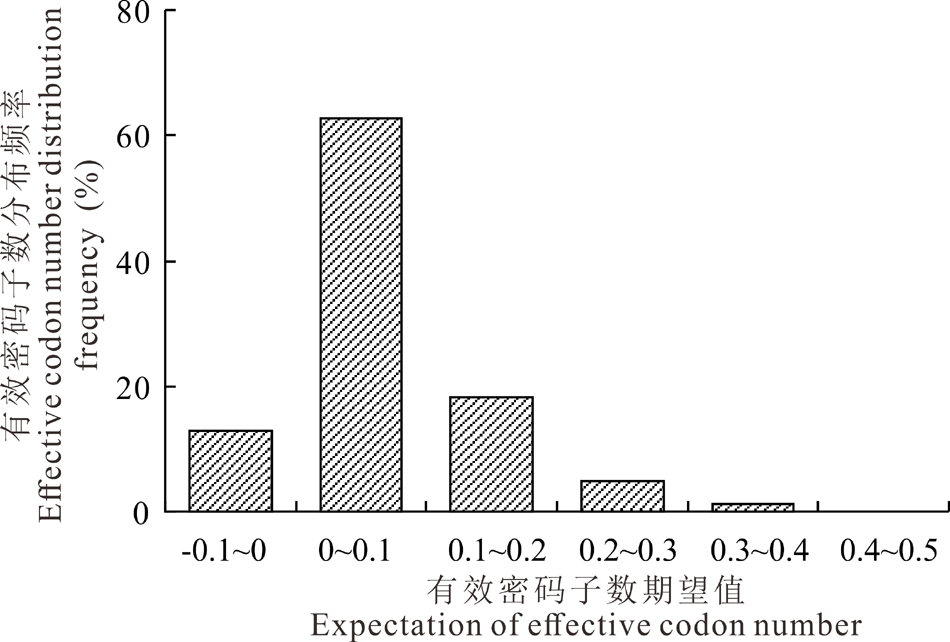 | 图2 内生真菌E. gansuensis基因组有效密码子数(ENC)的频数分布Fig.2 Frequency distribution of ENC on E. gansuensis |
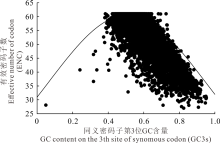
通常用ENC绘图分析来探讨GC3s对基因组密码子使用模式的影响。如果一个基因的有效密码子数(ENC)位于预期的曲线上, 则说明该基因对密码子的使用没有偏好性。本研究中, 大多数有效密码子数低于期望值且位于期望曲线的右下方(图3)。除了其他因素的影响, 突变压力也是影响其密码子使用偏好性的一个因素。
 | 图3 甘肃内生真菌基因组有效密码子数绘图分析ENC与GC3s之间的相关性Fig.3 Effective number of codons ENC-plot showing relationship between ENC and GC3s of E. gansuensis |
为了确定甘肃内生真菌基因组相对密码子使用偏好性和核苷酸组成之间的相关性, 本研究中使用了多变量的相关性分析对密码子使用偏好性和疏水性、芳香性以及基因长度之间的相互关系进行了确定(表2)。结果显示, 该基因组中, GC3s与Gravy、Aromo值均呈极显著相关(P< 0.01)。ENC值与Aromo(r=-0.022, P< 0.05)、Gravy(r=-0.159, P< 0.01)呈显著或极显著负相关, 这表明Aromo和Gravy值与肃内生真菌基因组密码子的偏好性呈显著的负相关。ENC值与基因长度呈极显著正相关(r=0.200, P< 0.01), 这说明基因的长度对密码子的偏好性使用有正向的帮助。同时, ENC值分别与第一轴(r=-0.876, P< 0.01)呈极显著负相关、与第二轴(r=0.108, P< 0.01)呈显著正相关、与GC3s呈极显著负相关(r=-0.808, P< 0.01)。
| 表2 各参数的相关性分析 Table 2 Correlation analysis of parameters |
分析结果表明, 在甘肃内生真菌基因组中CAI分别与GC (r=0.076, P< 0.01)、GC3 (r=0.265, P< 0.01)、GC3s (r=0.269, P< 0.01)、T3s (r=0.117, P< 0.01)和C3s (r=0.540, P< 0.01)均呈极显著正相关, 与GC1 (r=-0.043, P< 0.01)、GC2 (r=-0.244, P< 0.01)、A3s (r=-0.495, P< 0.01)、G3s (r=-0.190, P< 0.01)呈极显著负相关。此外, CAI分别与Gravy (r=0.086, P< 0.01)、Aromo (r=0.147, P< 0.01)呈极显著正相关。这些结果都表明, 甘肃内生真菌基因组对密码子使用的偏好性同样受到核苷酸组成和基因表达水平的影响。
运用统计学方法对CAI和密码子使用偏好性之间的关系进行分析, 并通过氨基酸相关指标分别沿4个向量轴对各基因进行相关性分析(表3)。
| 表3 对所有编码氨基酸的基因沿4个向量轴对各个位置的基因进行相关性分析 Table 3 Correlation coefficients between the positions of genes along the first four major axes with an index of total genes’ amino acid usage |
在甘肃内生真菌基因组中, CAI值除与第二轴(r=-0.590, P< 0.01)呈极显著负相关之外, 与其他指标之间均呈极显著正相关(P< 0.01)。CAI与Gravy (r=0.086, P< 0.01)和Aromo (r=0.147, P< 0.01)均呈极显著正相关。
根据分析得出, 甘肃内生真菌基因组所有最优密码子如表4所示。表中显示的是编码每个氨基酸的同义密码子, 以及每个同义密码子对应的“ 高表达” 和“ 低表达” RSCU值以及该密码子被使用过的次数。在甘肃内生真菌基因组中总共鉴定到26个最优密码子, 其中以G结尾的10个、以C结尾的16个, 表明供试基因组偏好使用的密码子可能与密码子第3位的GC含量有关。此外, 甘肃内生真菌编码精氨酸(Arg)的最优密码子有3个, 分别为AGG、CGC和CGG, 编码丝氨酸(Ser)的最优密码子也有3个, 分别为UCC、UCG和AGC, 编码丙氨酸(Ala)、苏氨酸(Thr)、脯氨酸(Pro)和亮氨酸(Leu)的最优密码子分别也有2个。
| 表4 内生真菌E. gansuensis基因组的最优密码子确定 Table 4 Optimal codons of genes in E. gansuensis |
基因的密码子偏好性是分析物种进化的一个重要参数, 并且在众多原核和真核生物研究中得到了证实。这些研究中得到了两个重要的结论, 即:中性进化和自然选择是影响物种密码子使用偏好性的最主要因素, 而且这两个结论已经被普遍用来解释形成不同物种密码子使用偏好性的起源[3, 36, 37]。当同义突变发生在密码子第3位上时, 将导致一个随机的密码子被选择使用, 一个基因中兼并密码子的碱基GC和AT被替代的比率是相同的, 但是这并不会改变翻译出的氨基酸种类[27, 38]。与此相反, 如果选择翻译的压力影响密码子的使用, 则说明密码子使用的偏好性与该基因的表达水平呈显著的正相关, 那么一些翻译偏好使用的密码子出现的频率相比其他一些将会更高。前人研究证实, 同一物种的基因常会分享相似的密码子使用模型。但是也有例外, 例如在枯草芽孢杆菌(Bacillus subtilis)的不同基因上密码子的使用模型有所差异[39]。
有研究报道称, GC3s分布的广度可能与定向选择的变化相关, 而与突变压力的相关性不强[32, 40]。甘肃内生真菌基因组中, GC3的分布范围主要集中在40%~60%, 分布广度相对较小, 这表明该菌对密码子的选择可能受到一定突变压力的影响。ENC绘图分析是基于GC3s对比分析某些基因实际观测值和期望值之间的分布关系, 能够揭示密码子使用偏好性是否受到突变的影响[27, 41]。本研究中, 大多数基因的有效密码子数位于期望值曲线的右下方, 低于期望值(图3)。表明除了其他因素的影响, 突变压力也是影响该菌密码子使用偏好性的一个重要因素。在对其他禾草内生真菌基因组密码子使用模式的研究中也得到了较类似的结果[42, 43], 突变压力在一定程度上对该菌密码子使用的选择具有一定的影响。
本研究中突变对密码子使用偏好性的影响仅为27.29%, 这意味着除中性进化之外的其他因素对甘肃内生真菌密码子使用的偏好性影响所占比例约为82.71%。此外, GC12和GC3之间呈极显著相关(P< 0.01), 且其回归系数为1.110。同时, 在中性分析中二者呈显著相关(P< 0.01), 这意味着对基因组中GC含量造成影响的主要因素是其内部GC的突变, 这个结果也可以用来解释密码子其他3个位置上的偏好性选择[32, 41, 44]。因此, 突变压力在禾草内生真菌密码子偏好性的选择上只扮演着很微小的作用, 而自然选择在其中可能扮演着重要的主导作用[42, 43]。
通过分析密码子使用的各指标与两类氨基酸之间的相关性, 结果表明疏水性氨基酸对甘肃内生真菌密码子使用偏好性的影响最大, 其次是芳香族氨基酸。这一结论为氨基酸的翻译效率是影响其密码子使用偏好性的重要因素提供了较强有力的证据[32, 40, 43], RNA的翻译效率也会影响密码子的使用频率。研究一致认为, 密码子的偏好程度与对应的基因表达水平具有密切联系, 高表达基因比低表达基因表现出更强的密码子使用偏好性。因此, 在自然选择压力下, 高表达基因偏好使用最优密码子[41, 45, 46, 47]。此外, 尽管本研究对一些特殊氨基酸同义密码子使用模式的相关指标进行了相关性分析, 但是有关氨基酸组成等因素与密码子使用模式之间的相关性仍需要进一步的深入研究。有学者报道称某些研究忽视了基因组中氨基酸的组成。例如, 在对基因组密码子使用模式的研究中, 遗漏平行加权, 造成分析结果被低估, 同时也会导致翻译错误[48]。
本研究发现, 甘肃内生真菌含有28个被高频使用的密码子, 密码子使用的偏好性较弱。另外, 这28个被高频使用的密码子均以G或C结尾。基于“ Δ RSCU” 指标对供试基因组的最优密码子进行鉴定, 共鉴定到26个最优密码子, 其中以C结尾的最优密码子16个、以G结尾的10个, 表明甘肃内生真菌偏好使用的密码子可能与第3位的GC含量有关。这一结果与其他禾草内生真菌基因组密码子使用偏好性的结果较为一致, 这类真菌对密码子使用的偏好性均较弱, 且最优密码子大多以G或C结尾, 但与其他类群的真菌或植物对密码子使用的偏好性差异较大[40, 41, 44]。对水稻(Oryza sativa)[33]、小麦(Triticum aestivum)[32]、玉米(Zea mays)[49]、蒺藜苜蓿(Medicago truncatula)[50]等一些高等植物核基因组的研究发现, 它们的最优密码子与禾草内生真菌的最优密码子较为类似, 均是以G或者C结尾。但是甘肃内生真菌核基因组最优密码子的特点与大肠杆菌、枯草芽胞杆菌、网柄菌(Dictyostelium discoideum)、黑腹果蝇、裂殖酵母(Schizosaccharomyces pombe)、酿酒酵母等其他酵母菌(Saccharomyces spp.)却明显不同[7, 51]。本研究中, 接近1/3的最优密码子是以尿嘧啶(U)结尾, 而其他的是以胞嘧啶(C)或鸟嘌呤(G)结尾, 这可能与他们的进化起源及亲缘关系相关。此外, 甘肃内生真菌编码精氨酸(Arg)和丝氨酸(Ser)的最优密码子有3个, 编码丙氨酸(Ala)、苏氨酸(Thr)、脯氨酸(Pro)和亮氨酸(Leu)的最优密码子分别也有2个。这些最优密码子可能与转录水平具有较强的相关性。
总之, 通过对甘肃内生真菌核基因组密码子使用模式的分析发现, 其密码子使用的偏好性均受到核苷酸的组成、突变压力、中性选择和基因表达水平的影响, 但影响程度较弱。然而, 自然选择在塑造密码子偏好性使用过程中扮演着非常重要的作用, 从而维持这种内生真菌相对较弱的密码子偏好性。此外, 相对于拟南芥、大肠杆菌或秀丽隐杆线虫(Caenorhabditis elegans)等模式物种[34], 本研究中所涉及的真菌对密码子的使用具有更大的偏好性。
随着越来越多的禾草内生真菌核基因组和线粒体基因组被组装完成并完全释放, 在未来对同一物种的核基因组与线粒体基因组密码子使用的对比分析将成为可能, 进而从另一个侧面揭示禾草内生真菌的进化起源。此外, 由于禾草内生真菌作为一种特异性较强的共生微生物, 不同菌株之间具有较强的宿主特异性, 是否可以通过对比分析某一宿主特异性较强的内生真菌及其宿主禾草的密码子偏好性, 进而在一定程度上揭示二者的协同进化机制。同时也可以对比分析分离自同一宿主植物不同种内生真菌或不同种群的同种内生真菌密码子使用的偏好性, 解释内生真菌基因型对宿主禾草的选择性机制或环境适应性机制。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|