{kind=link}
{kind=link}
{kind=link}
{kind=link}
甘草属杂交区物种形态特征的数值分类学研究
[陈雯清1  , 陆嘉惠
, 陆嘉惠1, 2, * , 王倩倩1 , 辛倩1 , 徐影1 ]
, 陆嘉惠, 王倩倩|
|


作者简介:陈雯清(1994-),女,河南信阳人,在读硕士。E-mail: 1607749254@qq.com
本研究对来自新疆和甘肃9个甘草属杂交区物种的形态分类性状进行数值分类分析,以区别和鉴定不同杂交区的亲本种组成及杂交类型,为药用甘草种质资源利用和杂交物种形成研究奠定形态分类学基础。通过野外调查和标本采集,对形态学特征及数据进行记录和测量;数值化处理后,进行主成分分析,提取具有分类价值的主要特征性状;通过R聚类对29个性状进行分类;通过OTU散点和Q聚类对不同杂交区的亲本种及其杂交类型进行分类。结果表明:在前3个主成分中果荚膨胀度(LeI)、小叶数上限(ULF)、小叶数下限(LLF)、果荚被腺体毛(LeG)、复叶长(CLL)、每荚含种子数(LePS)、复叶轴长(CRL)、小叶叶缘(LM)、果穗长/果穗宽(EL/EW)、果穗长(EL)、小叶长/小叶宽(LL/LW)、小叶宽(LW)的绝对权重值均在0.7以上,这12个性状在甘草属植物的分类学研究中具有重要价值。29个性状被归为2类:A类主要为叶片和果穗相关的性状;B类主要为果荚和复叶相关的性状,两类性状间的相关系数皆为1,表明这些性状相伴发生,具有很高的相关性。344份材料被分为A类胀果甘草;B类胀果甘草和乌拉尔甘草的杂交类型;C类乌拉尔甘草;D类光果甘草;E类胀果甘草和光果甘草的杂交类型。A类胀果甘草包括新疆阿拉尔、库尔勒和甘肃瓜州、民勤的胀果甘草,聚为一支,表明胀果甘草的不同居群遗传分化较小,形态特征不存在地域分布差异;C类甘肃民勤、张掖和新疆库尔勒、焉耆、博湖的乌拉尔甘草,分别被聚为两支,表明乌拉尔甘草出现种内遗传分化,甘肃和新疆不同分布区存在明显地域差异;D类光果甘草包括新疆阿拉尔的密腺甘草和光果甘草,支持密腺甘草为光果甘草的变种。9个自然杂交区类型分为3种:新疆阿拉尔、吐鲁番为胀果甘草与光果甘草的种间杂交区;新疆焉耆、库尔勒和甘肃瓜州为胀果甘草与乌拉尔甘草的种间杂交区;鄯善为胀果甘草、光果甘草及乌拉尔甘草的混合杂交区。所有杂交区的杂交类型均具有不同的种间镶嵌式形态特征。
In this study, the morphological characters of Glycyrrhiza specimens from nine regions of Xinjiang and Gansu Provinces where natural hybrids occur were classified by numerical taxonomic study to identify the parent species and hybridization type in different geographic areas where hybrids occur. Following field investigation and specimen collection, data on morphological traits were measured and recorded. After the numerical data collection, principal component analysis was carried out to identify the most important traits for classification purposes, and 29 traits were evaluated by R-cluster. The parental species and their hybrids in the different areas of hybrid occurrence were classified by operational taxonomic unit scatter and Q clustering. Principal component analysis result shows that Among the first three principal components, degree of legume inflation, upper limit of foliolate , lower limit of foliolate, presence of legume glandular hair, compound leaf length, no. of seeds per pod, compound rachis length, leaflet margin morphology, infructescence length, infructescence length:width ratio, leaflet length:leaflet width ratio, and leaflet width all are above 0.7, meaning that these twelve characters play an important role in taxonomic study on Glycyrrhiza species. The 29 traits could be classified into 2 categories: the first, mainly comprising leaflet and inflorescence traits, and the second, mainly comprising legume and compound leaf traits. The correlation coefficient between scores of the two categories of traits was 1, indicating that these trait groupings occur in conjunction and have high relevance as taxonomic indicators. Glycyrrhiza accessions ( n=344) were classified into groups as follows: A, Glycyrrhiza inflata; B, The hybrid of G. inflata and Glycyrrhiza uralensis; C, G. uralensis; D, Glycyrrhiza glabra; E, the hybrid of G. inflata and G. glabra. A variation of G. inflata includes plants from Alaer and Korla in Xinjiang, and Guazhou and Minqin in Gansu Provinces, which are gathered into one branch, as these populations of G. inflata are less genetically differentiated. Plants of group C from Minqin, Zhangye in Gansu and from Korla, Yanqi, and Bohu in Xinjiang, were divided into two branches, as intravarietal genetic differentiation had occurred. Group D representing G. glabra. includes G. glabra and Glycyrrhiza glabra var. glandulosa from Alaer, Xinjiang, as G. glabra var. glandulosa is a variety of G. glabra. Nine geographic areas where natural interspecific hybrids occur could be divided into three categories: Alar and Turpan in Xinjiang Province are interspecific hybridization areas of G. inflata and G. glabra; Yanqi and Korla in Xinjiang and Guazhou in Gansu are interspecific hybridization areas of G. inflata and G. uralensis; Shanshan in Xinjiang is a mixed hybridization area of G. inflata, G. glabra and G. uralensis. All hybrid specimens of Glycyrrhiza examined, have distinctive and different inter-species combinations of morphological features. This study laid the foundation of morphological taxonomy for the utilization of medicinal Glycyrrhiza germplasm resources and the formation ofhybrid species.
豆科(Leguminosae)甘草属(Glycyrrhiza)是蝶形花亚科山羊豆族甘草亚族的唯一一个属[1], 属内植物为多年生草本或半灌木; 在全世界总计有21种, 主要分布于中亚、北美及东欧等地; 我国有8种, 主要分布于黑龙江、吉林, 内蒙古、新疆、甘肃等地[2]。其中我国分布的3种甘草属植物, 胀果甘草 (Glycyrrhiza inflate)、乌拉尔甘草(Glycyrrhiza uralensis)和光果甘草(Glycyrrhiza glabra)为“ 甘草” 基原植物被收录于《中华人民共和国药典(第一部)》[3]。
杂交区是指不同基因型的种群相遇, 交配并产生杂交后代的狭窄地带[4, 5]。不同的杂交带中生长着不同的亲本物种以及没有独立遗传世系的近缘种[6], 在长期的自然演化中, 这些物种相互竞争并不断进行形态分化以及传粉、繁殖等机制的调整以适应自然环境[7]。最新的研究和野外调查发现, 甘草属在自然生境中存在属内不同物种的同域分布区, 且种间亲和性高, 极易形成自然“ 杂交区” 或“ 杂交带” [8]。杂交区内, 甘草属亲本及其杂交种长期混居在一起, 种间杂交及不同程度和方向地回交导致的杂交渐渗使得杂交区产生了不同的种内变异类型及种间过渡类型[9, 10, 11]。在外部生境的自然选择压力和内部的杂交基因渐渗共同作用下, 这些类型在形态特征上通常表现为其叶、花部、果部各性状的连续变化或者间断变化[12, 13]。
目前, 由于对甘草属杂交区亲本和杂交类群形态分化没有明确的分类依据和认识, 往往把杂交类群也当成了亲本种, 造成了药用甘草资源应用和研究的混乱, 种源不清, 人工栽培引种后幼苗形态出现分化, 严重影响甘草药材生产质量管理规范(GAP)的进程[14, 15]。同时, 由于对甘草属自然杂交区缺乏比较全面、系统的野外调查和对杂交区物种的适应进化研究, 在理论和实践应用方面, 也无法对出现的分类有争议的野生甘草资源澄清“ 身份” , 明确类型。
本研究通过野外调查、标本采集、形态性状的数值分类方法, 对甘草属9个自然杂交区的344份材料的主要分类性状和特征进行数值分类, 分析不同杂交区的亲本种组成及杂交类型的差异, 揭示甘草属不同杂交区亲本种及其杂交类型, 为药用甘草种质资源利用和杂交物种形成机制奠定形态分类学基础。
于2018年9-10月对分布于新疆、甘肃的甘草属自然杂交区进行野外调查和标本采集, 用于数值分类研究的标本数为344份。主要为胀果甘草、乌拉尔甘草和光果甘草及其杂交后代, 采集地详见表1。
| 表1 甘草属9个杂交区试验材料 Table 1 The material of 9 hybrid zones of Glycyrrhiza |
1.2.1 性状选取、测定及编码 对9个自然杂交区的甘草属植物进行形态性状分析, 选取29个表型性状进行编码, 具体性状描述及编码参考Agrimonti等[12]、Hassannejad等[13]、张富民等[16]的研究。其中质量性状11个, 数量性状14个, 比例指标4个, 定性性状编码赋值由弱至强, 由少至多, 具体编码和数值见表2。
| 表2 基于形态学性状的甘草属植物的性状编码 Table 2 Character coding of Glycyrrhiza based on morphological characters |
1.2.2 数量性状的测定 小叶数、复叶长、叶轴长、小叶长和宽、叶柄长、小叶最宽处至叶尖长等叶片形态数据为每份标本上随机抽取10个复叶进行测量, 取平均值; 果穗长、宽、花序轴长、果荚长、宽、厚、每荚含种子数等果的形态数据为每份标本上取所有果穗进行测量, 计算平均值, 所有测量值长度单位均为cm。每种类型的植物测定样本数详见表1。
1.2.3 数据分析 采用R语言计算各性状间相关系数得相关系数矩阵; 根据成分提取结果确定主成分, 求出各主成分的特征值、贡献率、累计贡献率和主要因子; 以主成分1、2为横、纵坐标作运算的分类单位(OUT)散点图。采用SPSS软件对标本进行聚类分析, 运算时对原始数据进行标准差标准化处理; 采用平均欧氏距离平方系数作Q聚类, 分析各类群甘草的性状相似度; 采用皮尔逊相关系数作R聚类, 分析各性状之间的相关性。
2.1.1 主成分提取 通过SPSS软件对344份标本的29个性状进行了主成分分析, 提取了特征值大于1的7个主成分。由表3可知, 前7个主成分的累计贡献率为77.308%。第1主成分所占信息达28.494%, 第2主成分所占信息达17.573%, 第3个主成分所占信息达11.420%, 第4个主成分所占信息达6.616%, 第5个主成分所占信息达5.175%, 第6个主成分所占信息达4.460%, 第7个主成分所占信息达3.570%, 而其余22个成分所提供的信息量仅为22.390%。表明在甘草分类研究中存在重要性状, 有必要做主成分分析。
| 表3 甘草属不同形态性状的主成分分析 Table 3 Principal component analysis of different morphological traits in Glycyrrhiza |
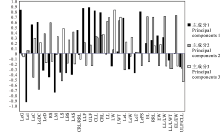
2.1.2 性状的主成分载荷分析 通过成分矩阵, 根据性状在主成分中所占的总载荷量, 将初始因子载荷矩阵和性状抽提系数表综合起来, 对各性状进行比较, 得到甘草属29个性状特征对应的主成分载荷量和主要因子。由图1可知, 主成分1中, 果荚膨胀度(LeI)、小叶数上限(ULF)、小叶数下限(LLF)、果荚被腺体毛(LeG)、复叶长(CLL)、每荚含种子数(LePS)、复叶轴长(CRL)、小叶叶缘(LM)8个性状均具有较高的载荷量, 绝对值在0.7以上; 第2主成分中, 果穗长/果穗宽(EL/EW)、果穗长(EL)、小叶长/小叶宽(LL/LW)3个性状占有较高的载荷量, 绝对值大于0.7; 第3主成分中, 小叶宽(LW)占有较高的载荷量, 绝对值为0.84。表明这些性状为前3个主成分的主要构成因子, 在甘草属植物的分类学研究中具有重要意义。
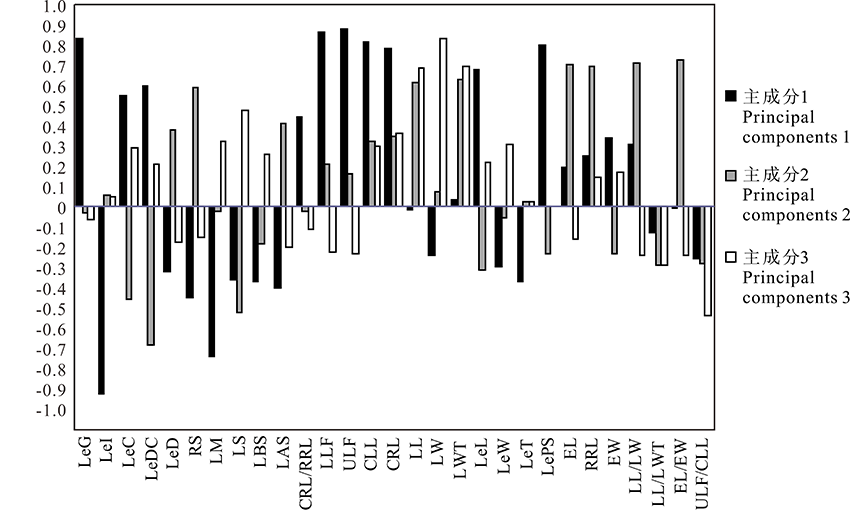 | 图1 甘草属29个性状前3个主成分的因子载荷矩阵Fig.1 Factor loading matrix of the first three principal components of 29 characters of Glycyrrhiza |
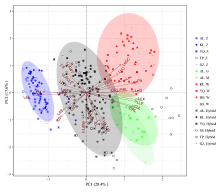
2.1.3 基于主成分分析的OTU散点图分类 由图2可知, OTU散点图将344份甘草分为5种类型:胀果甘草分布在二、三象限; 乌拉尔甘草主要分布在第一象限; 光果甘草和密腺甘草分布在第四象限; 杂交类型以坐标原点为中心, 散布于4个象限中, 但与3个亲本种很少重叠, 根据与亲本种的距离远近, 可以判断第一、二象限的为胀果甘草与乌拉尔甘草的杂交类型; 第三、四象限的为胀果甘草与光果(密腺)甘草的杂交类型。
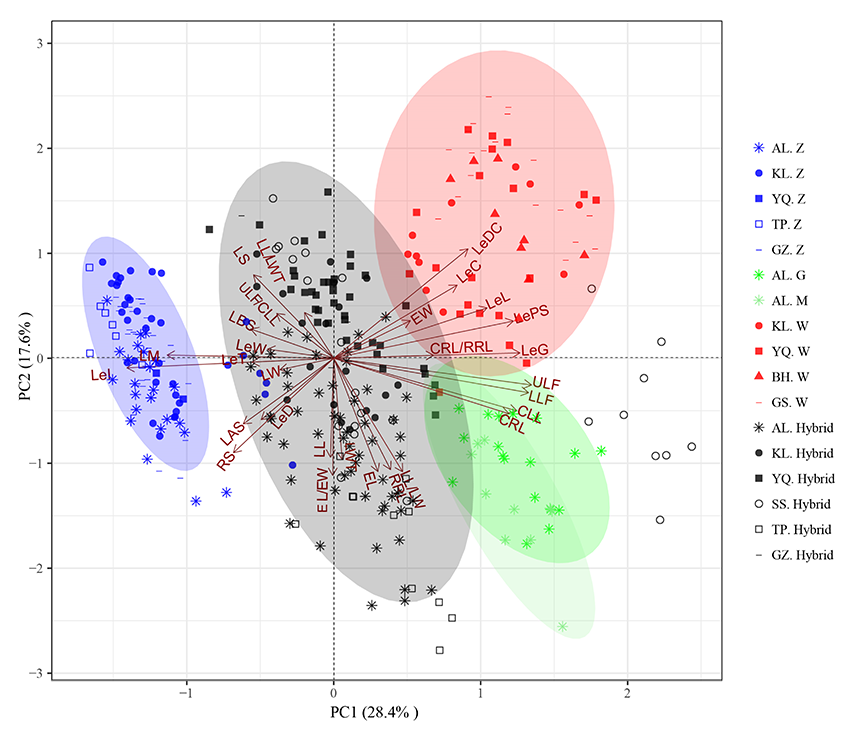 | 图2 基于第一、二主成分的344份甘草属植物OTU散点图Fig.2 OTU scatter plots of 344 Glycyrrhiza based on the first and second principal components |
果荚膨胀度(LeI)、小叶数上限(ULF)、小叶数下限(LLF)、果荚被腺体毛(LeG)、复叶长(CLL)、每荚含种子数(LePS)、复叶轴长(CRL)、小叶叶缘(LM)8个性状与X轴的余弦值较大, 在第一主成分中占比较高, 这些性状将甘草属分为3类:胀果甘草为一类, 乌拉尔甘草和光果甘草(含密腺甘草)为一类, 3种甘草的杂交类群为一类。由OTU散点图中射线所表达的相关性可知, 果荚膨胀度(LeI)、小叶叶缘(LM)这两个性状与胀果甘草成正相关, 与乌拉尔甘草和光果甘草呈负相关; 每荚含种子数(LePS)、果荚被腺体毛(LeG)、小叶数上限(ULF)、小叶数下限(LLF)、复叶长(CLL)、复叶轴长(CRL)这6个性状与乌拉尔甘草和光果甘草呈正相关, 与胀果甘草呈负相关。结合形态特征分析, 胀果甘草具有果荚膨胀度高、小叶叶缘皱、每荚含种子数少、果荚不被腺体毛、小叶数少、叶轴短的特征; 乌拉尔甘草和光果甘草具有果荚膨胀度低, 叶缘不皱或微皱、每荚含种子数多、果荚被腺体毛、小叶数多、叶轴较长的特征; 杂交类群的这些性状均介于亲本种中间。果穗长/果穗宽(EL/EW)、果穗长(EL)、小叶长/小叶宽(LL/LW)这3个性状与Y轴的余弦值较大, 在第二主成分中占比较高, 这些性状不仅显著将甘草属2个亲本种乌拉尔甘草、光果甘草的类群分开, 同时也将不同杂交类群分为2类:第一、二象限的为胀果甘草与乌拉尔甘草的杂交类型; 第三、四象限的为胀果甘草与光果(密腺)甘草的杂交类型。由OTU散点图中射线所表达的相关性可知, 果穗长/果穗宽(EL/EW)、果穗长(EL)、小叶长/小叶宽(LL/LW)与光果甘草、胀果甘草同光果甘草的种间杂交类群成正相关, 与乌拉尔甘草、胀果甘草同乌拉尔甘草的种间杂交类群呈负相关。结合形态特征分析, 光果甘草具有果穗长/果穗宽比值大、果穗长, 小叶长/小叶宽比值大的特征; 乌拉尔甘草果穗长/果穗宽比值小、果穗短, 小叶长/小叶宽比值小; 杂交类群的这些性状均介于亲本性状中间。由以上分析可知, 仅用两个主成分和提取的主要性状就可以将各亲本种和杂交类群进行区分, 表明基于形态性状的主成分分析方法对甘草属物种分类学研究是可行的。
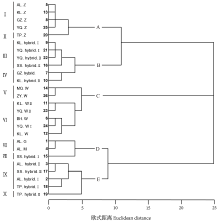
Q聚类中, 甘草属植物被归为5个类型:A为胀果甘草, 分别为Ⅰ 类群阿拉尔、库尔勒、瓜州、焉耆的胀果甘草, Ⅱ 类群吐鲁番的胀果甘草。B为胀果甘草和乌拉尔甘草的杂交类型, 分别为Ⅲ 类群瓜州、焉耆、库尔勒、鄯善的杂交类型, Ⅳ 类群库尔勒杂交类型。C为乌拉尔甘草, 分别为Ⅴ 类群民勤和张掖的乌拉尔甘草, Ⅵ 类群库尔勒、焉耆、博湖的乌拉尔甘草。D为光果甘草及其变异类型, 分别为Ⅶ 类群阿拉尔的光果甘草和密腺甘草, Ⅷ 类群鄯善变异类型。E为胀果甘草和光果甘草的杂交类型, 分别为Ⅸ 类群阿拉尔、吐鲁番、鄯善的杂交类型, Ⅹ 类群阿拉尔的杂交类型(图3)。
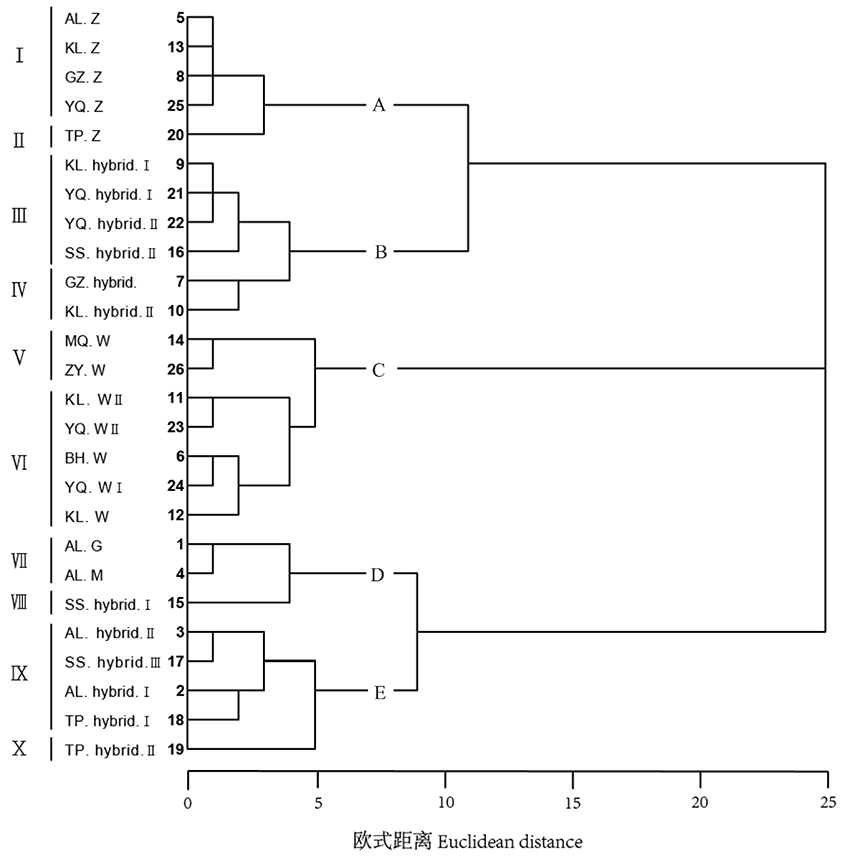 | 图3 基于29个形态性状的甘草属植物的亲缘关系树状图Fig.3 Genetic relationship treeplot of Glycyrrhiza based on 29 morphology characters |
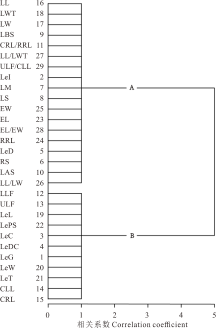
R聚类中, 29个性状被归为2类, A类主要为叶片和果穗相关的性状, 包括小叶长(LL)、小叶最宽处至叶尖长(LWT)、小叶宽(LW)等18个性状, 相关系数皆为1, 表明这些性状相伴发生, 具有更高的相关性; B类主要为果荚和复叶相关的性状, 包括小叶数下限(LLF)、小叶数上限(ULF)、荚果长(LeL)等11个性状, 相关系数皆为1, 表明这些性状相伴发生, 具有更高的相关性(图4)。
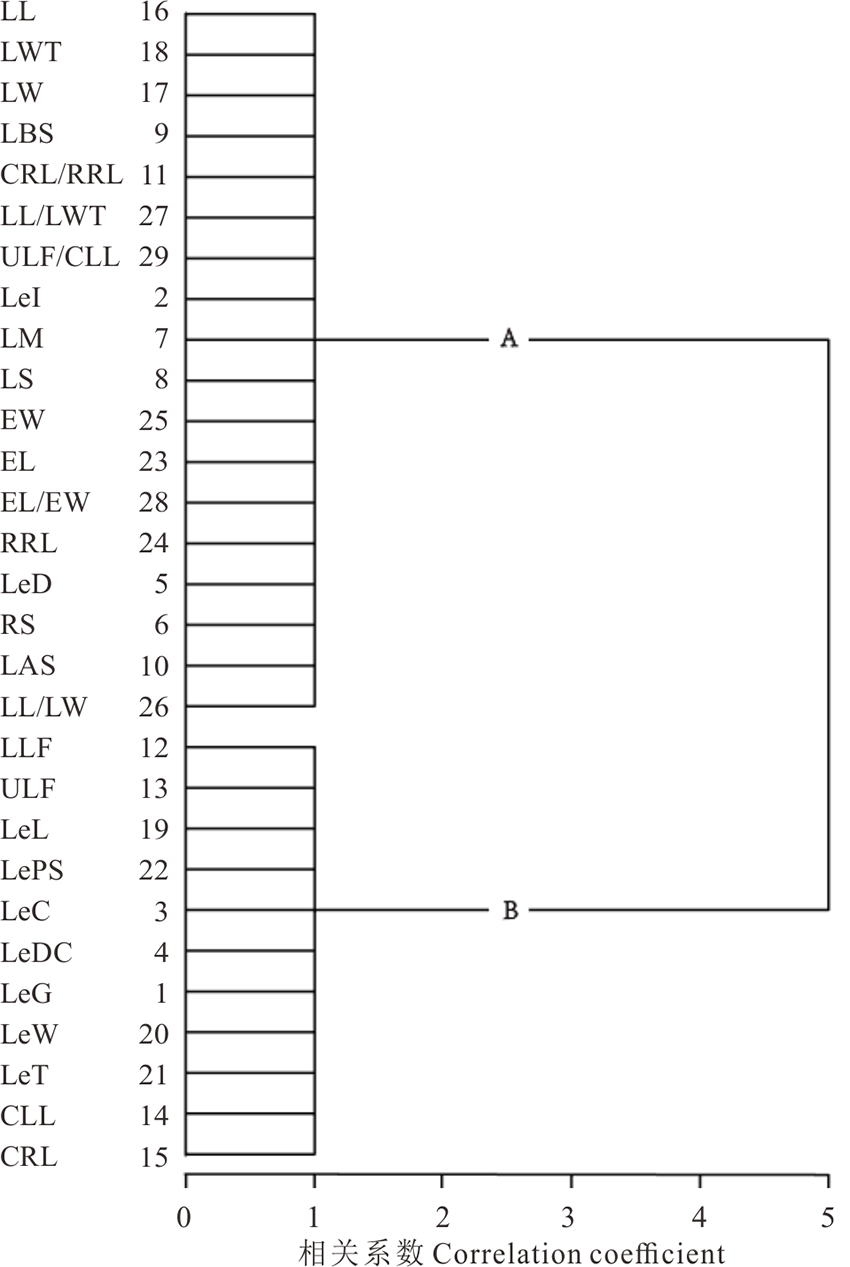 | 图4 甘草属植物29个形态性状的聚类图Fig.4 Cluster analysis of 29 morphological traits of Glycyrrhiza |
结合OTU散点图和聚类分析结果, 新疆和甘肃的甘草属自然杂交区的亲本种组成及杂交类型为:新疆阿拉尔、吐鲁番为胀果甘草与光果甘草的种间杂交区, 分布有胀果甘草、光果甘草、密腺甘草以及两种种间杂交类型; 新疆焉耆、库尔勒、甘肃瓜州为胀果甘草和乌拉尔甘草杂交区, 分布有胀果甘草、乌拉尔甘草以及两种种间杂交类型; 新疆鄯善为胀果甘草、光果甘草及乌拉尔甘草的混合杂交区, 在聚类分析和基于主成分分析的散点图中, 其杂交类型分散在胀果甘草、乌拉尔甘草、光果甘草类群及其种间杂交类群中, 因此其物种成分比较复杂。
同一杂交区内的杂交类型存在不同程度地分化:在胀果甘草与乌拉尔甘草的种间杂交区中, 存在Ⅲ 、Ⅳ 两种杂交类群, 其果荚和小叶数、叶形往往兼具了2个亲本的特点, 表现为果荚微胀、小叶微皱、小叶数均为5~9, 介于2个亲本的范围内。但两种杂交类群又具有一定的分化, 例如, Ⅲ 类群果荚更为膨胀且果荚直, 与胀果甘草形态更为相近; Ⅳ 类群果荚微弯, 种子间有起伏凹凸, 叶片形态更接近乌拉尔甘草(表4)。在胀果甘草与光果甘草的种间杂交区, 存在Ⅷ 、Ⅸ 、Ⅹ 3种杂交类群, Ⅷ 类群果荚、复叶形态特征与光果甘草最为相似, 且小叶数明显多于光果甘草, 可能为光果甘草的变异类群(表5); Ⅸ 类群和Ⅹ 类群果荚和小叶数兼具了两个亲本的特点, 果荚长度居于两亲本之间, 小叶数均为5~11, 其中Ⅸ 类群果荚稍长微弯, 形态特征与光果甘草更为相近; Ⅹ 类群果荚较短且直, 与胀果甘草更为相近。此外, 密腺甘草和光果甘草聚为一支, 形态相似; 阿拉尔、库尔勒、瓜州、焉耆分布的胀果甘草和吐鲁番的胀果甘草聚为两支, 但形态特征分化较小; 而甘肃和新疆分布的乌拉尔甘草聚为两支, 形态差异较为明显。
| 表4 乌拉尔甘草、胀果甘草及其杂交类型的形态学比较 Table 4 Morphological comparison of G. uralensis, G. inflate and their hybrid types |
| 表5 胀果甘草、光果甘草及其杂交类型的形态学比较 Table 5 Morphological comparison of G. inflate, G. glabra and their hybrid types |
主成分提取可以将多个形态特征提取为几个主成分因子, 从而达到降维的目的。目前, 形态学和分子证据进行的主成分分析已成功用于证明不同动植物的杂交和基因渐渗[17, 18, 19]。甘草属果荚和叶片的形态特征一般在种内通常较为稳定, 是鉴定3种药用甘草的主要分类性状[16, 20]。本研究通过对甘草属果荚和叶片的形态特征进行分析, 对3种药用甘草及其杂交类群的主要性状进行提取, 提取出果荚膨胀度(LeI)、小叶数上限(ULF)、小叶数下限(LLF)等12个形态性状为甘草属亲本种与其杂交类型的主要分类性状。
甘草属各个性状间存在很高的相关性, 表明相关性状可能在基因上存在某种连锁现象[21]。甘草属因子载荷矩阵显示, 随着果荚膨胀度越高, 小叶叶缘越皱, 每荚含种子数越少, 果荚被腺体毛越少, 小叶数越少, 叶轴越短; 小叶最宽处至叶尖长比值越大, 果穗越长, 小叶数上限/复叶长比值越大。这与Hassannejad等[13]的研究相符, 果荚被腺体毛程度, 每荚含种子数, 复叶轴长等具有极强的相关性。此外, 这些相关度高的性状, 通常突出表现亲本种的特征[12], 因此, 可以依据相关度高的性状特征识别甘草属的种类。
我国甘草种质资源变异丰富, 同种类不同居群间遗传分化大[22]。研究表明, 3种药用甘草在化学成分含量、生理生化指标, 生长环境等方面均有差异[23, 24, 25]。本研究从甘草属的外部形态特征对3种药用甘草进行分析, 聚类图和OTU散点图结果均将所有甘草属分为5类, 与《中国植物志》[1]的分类相吻合。在OTU散点图和聚类分析结果中, 新疆阿拉尔的密腺甘草和光果甘草形态上极为相近, 被聚为一类, 结合形态特征, 蜜腺甘草除了果荚密被腺体外, 与光果甘草形态基本一致, 支持密腺甘草作为光果甘草的变种处理, 这一结果与宋凤等[26]的研究相符。
新疆、甘肃分布的胀果甘草形态上极为相近, 其形态上均表现为果荚短小膨胀, 果内种子数少于4颗, 小叶数3~7, 在OTU散点图和聚类分析结果中, 除新疆吐鲁番的胀果甘草, 均被聚为一类, 表明胀果甘草在形态特征上不存在地域分布的差异, 但是否存在基因型的差异, 还有待进一步通过其他分类方法进行分析。另外, 胀果甘草种内形态分化小, 可能也与其相对狭窄的分布区有关。植物的遗传变异水平越低, 种群分布范围越小[27], 中国植物志记载及野外调查表明, 胀果甘草仅分布于中国新疆天山以南, 及向东延伸至甘肃酒泉等地, 其他地区再无分布[28], 胀果甘草的遗传变异水平较低。分布区范围往往也代表了物种形成和进化过程中对环境的适应能力, 根据文献记载, 胀果甘草是3种药用甘草中最喜光的[29], 也是耐旱、耐盐性最强的药用甘草[30, 31]。因此, 干旱、半干旱、光热资源丰富, 是胀果甘草分布的主要生境, 沿甘肃酒泉向东延伸, 由于受光热资源限制, 胀果甘草往往没有分布。
乌拉尔甘草适应幅度大且分布广, 在温带草原地区和暖温带荒漠地区中均有分布, 广泛出现于各地山区、河谷、杨树林及荒漠灌丛、低地河漫滩草地及灌溉绿洲[32]。地理阻隔和不同自然条件的长期作用可使形态特征在植物居群间产生较大的差异[33]。由于分布广泛, 空间地理上相距较远, 对生境的适应性进化, 乌拉尔甘草不同居群的遗传差异往往较大[34, 35]。本研究中, 乌拉尔甘草分别被分为两支, 代表了甘肃、新疆分布的乌拉尔甘草存在明显的地域分布差异, 主要表现为果序的紧实或松散、果荚背腹弯曲浅或弯曲深, 这与相关研究认为的乌拉尔甘草的种内变异类型多, 形态差别大, 种内遗传多样性高一致[36, 37, 38]。
杂交区中通常分布着许多的亲本种及杂交类群, 这些杂交类群有些是亲本与其杂交后代的关系, 有些属于近缘的“ 姐妹” [39, 40, 41], 它们同域分布且彼此间的隔离机制不完善, 在不同程度基因渐渗和外部环境因素的影响下, 其形态特征具有多样性[42, 43]。自然杂交种通常表现出其亲本物种镶嵌式的形态特征, 因此形态特征介于两亲本中间的物种通常可以判断杂交种, 但当存在一定程度和单一方向的基因渐渗时, 杂交种可能与其某一亲本物种极为相似[44]。本研究中胀果甘草与乌拉尔甘草的种间杂交类型, 其果荚, 小叶数, 叶形均兼具了2个亲本的特点, 但由于基因渐渗的不同, 在形态上又存在分化现象, Ⅲ 类群果荚形态与胀果甘草形态更为相近, 而Ⅳ 类群果荚形态更接近乌拉尔甘草, 2种种间杂交类型代表了不同变异方向; 胀果甘草与光果甘草的种间杂交类群Ⅷ 、Ⅸ 、Ⅹ 中, Ⅷ 类群果荚、复叶形态特征与光果甘草极为相似, Ⅸ 类群果荚形态特征与光果甘草更为相近, 而Ⅹ 类群果荚与胀果甘草更为相近。以上分析表明, 甘草属的自然杂交区内, 即使亲本种相同, 其后代在形态上也存在差异, 不仅仅表现为亲本种的中间性状, 有时也与亲本种既有相似的性状, 又有自己特有的杂交性状。这种杂交类型存在镶嵌式的形态特征, 往往给物种分类和杂交种鉴定带来困难, 因此在进行野外调查和采集时, 应注意调查地的生境和亲本物种类型, 以便对出现的种间镶嵌式的形态特征进行区别和分类。
杂交带的物种维持主要受内在选择和外在选择相互作用的影响, 内在选择是指杂交区亲本及其杂交后代基因型对杂交带生境的适合度的不同而导致杂交带物种的分化[45], 外在选择是指外在环境变化对杂交带物种的筛选[46]。另外, 种群的大小和数量也取决于其本身物种对生态因子的适应性或适合度[47, 48]。本研究中, 新疆和甘肃地区的自然杂交区, 同域分布种的种间杂交及杂交渐渗, 使杂交区产生了种间、种内的变异类型及亲本的中间过渡类型[49, 50], 这些杂交类群与其亲本种均能在同一生境中共存, 对生态因子有着很高的适合度。但与母体相比, 杂交类群是否存在适合度的分化, 或者出现以生殖隔离为标志的物种的分化, 还有待进一步研究。
近年来, 药用甘草已广泛应用于医药、发酵、饮料制作等方面, 野生药用甘草的需求量不断上升[51, 52, 53]。然而, 由于开荒、放牧、工业建筑以及无计划采挖等人为活动的影响, 野生甘草生境遭到严重的破坏[54]。我国甘草属资源分布广泛, 其中以乌拉尔甘草分布最广, 在华北、东北、江苏、内蒙古等省区均有分布[55, 56]。据记载, 新疆库尔勒、吐鲁番、鄯善等地均是野生乌拉尔甘草的自然分布区[2], 但在本次野外调查中, 这些分布区已很难找到种群集中、大片的分布, 往往只能在林下、农田生境中发现零星分布。这与乌拉尔甘草自然分布区往往是轻度盐碱生境有关, 适宜乌拉尔甘草生长的生境, 往往也成了开荒造田的首选地。因此, 农田开垦对乌拉尔甘草的生存环境造成了破坏, 人为扰动会对乌拉尔甘草造成消极影响[57], 对甘草属资源的保护刻不容缓。
通过对新疆、甘肃的9个甘草属自然分布区的亲本种及其杂交类型的数值分类研究, 结论如下:1) LeI(果荚膨胀度)、ULF(小叶数上限)、LLF(小叶数下限)、LeG(果荚被腺体毛)、CLL(复叶长)、LePS(每荚含种子数)、CRL(复叶轴长)、LM(小叶叶缘)、EL/EW(果穗长/果穗宽)、EL(果穗长)、LL/LW(小叶长/小叶宽)、LW(小叶宽)这12个分类性状是甘草属杂交类型与其亲本种的重要分类性状。2) 29个性状被归为2类:叶片和果穗相关的性状聚为一类, 果荚和复叶相关的性状聚为一类, 两类性状间的相关系数皆为1, 表明这些性状相伴发生, 具有很高的相关性。3) 新疆和甘肃分布的胀果甘草不存在地域分布差异, 但乌拉尔甘草出现种内遗传分化, 具有明显地域差异, 甘肃、新疆分为两支。4) 甘草属自然杂交区类型有3种:新疆阿拉尔、吐鲁番为胀果甘草与光果甘草的种间杂交区; 新疆焉耆、库尔勒和甘肃瓜州为胀果甘草与乌拉尔甘草的种间杂交区; 新疆鄯善为胀果甘草、光果甘草及乌拉尔甘草的混合杂交区; 杂交区、物种类型不同, 杂交类型也具有不同的种间镶嵌式的形态特征。本研究揭示了甘草属杂交区的组成及种质资源类型, 为药用甘草种质资源利用和杂交物种形成机制奠定形态分类学基础。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|