{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
19种菊科植物花药结构及绒毡层的发育类型研究
[王瑞 , 胡玥, 崔宏安, 牛佳睿, 李亚东, 苗芳
, 胡玥, 崔宏安, 牛佳睿, 李亚东, 苗芳* ]
, 胡玥, 崔宏安, 牛佳睿, 李亚东, 苗芳]
|
|


作者简介:王瑞(1998-),女,宁夏固原人,在读本科。E-mail: 1430438307@qq.com
花药结构和绒毡层的发育类型对菊科植物的系统分类和系统演化具有重要的意义。本研究采用苏木精和苯胺番红整体染色以及石蜡切片的方法,通过显微镜观察,对菊科7族19属19种植物花药结构及绒毡层的发育类型进行研究。结果表明,18种植物的花药由4个花粉囊组成,1种由2个花粉囊组成。花粉囊壁由表皮、药室内壁、中层和绒毡层4层细胞组成,中层细胞在花粉母细胞时期解体。随着花粉粒的形成,药室内壁先“U”形增厚,再进行条带状增厚。花粉母细胞胞质分裂类型有连续型和同时型2种,小孢子四分体以四面体形为主。绒毡层的发育类型有变形绒毡层和分泌绒毡层两种类型,变形绒毡层是管状花亚科植物的共同特点,分泌绒毡层是舌状花亚科植物的共同特点。从向日葵族向菊苣族的逐渐演化过程中,绒毡层的发育类型从向日葵族的变形绒毡层、经过变形绒毡层向分泌绒毡层的过渡类型逐渐演化为菊苣族的分泌绒毡层。变形绒毡层的形态在管状花亚科不同族之间有明显差异,绒毡层的发育类型和形态可以作为亚科和族分类的依据之一。
Anther structure and tapetum developmental type is important for systematic classification and understanding of the evolution of the Asteraceae. In this work, anther structures and tapetum development types of 19 species in 19 genera of the Asteraceae were investigated by paraffin sections, hematoxylin integral staining, aniline-saffron integral staining and microscopic observation. It was found that the anther structures of all species examined consist of four clinandria except for one containing two. The anther wall consists of four layers of cells, namely: the epidermis, endothecium, middle layer and the tapetum layer. The middle layer cells disintegrate in the meiocyte period. During formation of pollen grains, the endothecium thickens in a U-shaped manner initially, followed by band-shaped thickening. The microspore mother cells have two types of cytokinesis, a continuous type and a simultaneous type. The majority of the microspore tetrads are tetrahedral. The tapetum has two types, an amoeboid type, characteristic of the Carduoideae, and a glandular type, a feature of the Cichorioideae. From an evolutionary perspective, a progression of three tapetal developmental types is evident: the amoeboid tapetum of the Heliantheae, a transitional type from amoeboid tapetum to glandular tapetum and glandular tapetum in the Lactuceae. The morphologies of the amoeboid tapetum are obviously different among the various species. The developmental type and morphology of the tapetum are important characteristics in Asteraceae classification.
菊科(Asteraceae)是被子植物双子叶纲中最为进化的植物类群, 其种数和分布范围都居显花植物之最, 共 1479属, 21105种, 广布于世界各地[1]。菊科按照头状花序中小花的构造以及植物有无乳汁等特征, 分为舌状花亚科和管状花亚科, 舌状花亚科下分1个族, 管状花亚科下分12个族[2]。
80年代以来, 几个被子植物分类系统同时作了修订, 最重要的依据之一就是被子植物的胚胎学性状[3, 4, 5]。花的解剖结构受环境条件的影响相对较少, 因此, 保持了物种遗传上的稳定性, 在植物系统分类学上具有重要的参考价值。从花药横切面观察, 菊科植物花药通常由4个花粉囊组成, 花药壁的发育属于双子叶型, 花药壁由表皮、药室内壁、中层和绒毡层组成[6, 7, 8, 9, 10, 11, 12, 14, 15, 16, 17, 18, 19]。成熟的花药壁只有退化的表皮和纤维化增厚的药室内壁2层[6, 7, 8, 9, 10, 11, 12, 14, 15, 16, 19]; 也有表皮完全消失, 只保留1层纤维化增厚的药室内壁[18]; 有的成熟花药药室内壁不纤维化[17]; 有的花药没有中层, 只有表皮、药室内壁和绒毡层3层[13]。
根据绒毡层解体时是否离开原来的位置进入花粉囊, 绒毡层分为2种类型:变形绒毡层和分泌绒毡层。变形绒毡层在绒毡层解体时离开原来位置进入花粉囊中, 最终在花粉囊中解体消失。分泌绒毡层是绒毡层在原来的位置解体消失。据研究报道, 款冬(Tussilago farfara)[6]、台湾翅果菊(Pterocypsela formosana)[7]、小蓬草(Conyza canadensis)[8]和一年蓬(Erigeron annuus)[9]的绒毡层为分泌绒毡层; 革苞菊(Tugarinovia mongolica)[10]、黄顶菊(Flaveria bidentis)[11]、孔雀草(Tagetes patula)[12]、牛膝菊(Galinsona parviflora)[13]、三裂叶豚草(Ambrosia trifida )[14]、千里光(Senecio glaucus)[15]、金盏花(Calendula officinalis)[18] 、菊苣(Cichorium intybus)[19]和豚草(Ambrosia artemisiifolia)[16]为变形绒毡层。钻叶紫菀(Aster subulatus)[17]、马兰(Kalimeris indica)[17]、普陀狗娃花(Heteropappus arenarius)[17]和一年蓬[17]的绒毡层为分泌和变形绒毡层共存。
根据减数分裂胞质分裂的形态, 胞质分裂类型有两种:同时型和连续型。同时型是减数分裂的第1次分裂只进行核分裂而不进行质分裂, 等第2次核分裂完成后同时进行质分裂产生4个细胞, 即四分体。连续型是减数分裂第1次核分裂完成后即刻进行质分裂形成2个细胞, 即二分体, 二分体再进行第2次核分裂和质分裂形成4个细胞, 即四分体。据研究报道, 台湾翅果菊[7]、小蓬草[8]、一年蓬[9]、革苞菊[10]、金盏花[18]、钻叶紫菀[17]、马兰[17]、普陀狗娃花[17]均属于同时型。黄顶菊[11]、孔雀草[12]、牛膝菊[13]和豚草[16]均属于连续型。小孢子母细胞减数分裂形成四分体, 根据四分体的排列方式, 将四分体分为四面体形、左右对称形、十字形(交叉形)、直列形和T形5种形态。据研究报道, 台湾翅果菊[7]、革苞菊[10]、黄顶菊[11]、牛膝菊[13]、千里光[15]和豚草[16]的四分体为四面体形。小蓬草[8]的四分体具有四面体形和左右对称形2种。一年蓬[17]以四面体形为主, 兼有十字形和左右对称形。金盏花[18]的四分体兼有四面体形和十字形2种。
综上所述, 菊科植物花药壁的结构、绒毡层的类型、小孢子母细胞减数分裂胞质分裂类型、四分体的形态以及成熟花药壁的结构在不同种类之间具有较明显的差异。前人的研究主要集中在菊科某个种花药解剖结构的特征上, 还没有较系统地进行菊科亚科之间、族之间以及属间花药结构的比较研究, 该方面的研究对菊科植物的系统分类和进化具有重要的参考价值。本研究拟选择菊科管状花亚科向日葵族(Heliantheae)7个属、菜蓟族(Cynareae) 3个属, 春黄菊族(Anthemideae)、千里光族(Senecioneae)、堆心菊族(Helenieae)和旋覆花族(Inuleae)各1个属植物, 舌状花亚科(仅含1个族)菊苣族(Lactuceae) 5个属总计19种植物为材料, 观察比较亚科之间、族之间以及属间花药结构、绒毡层的类型、小孢子母细胞减数分裂胞质分裂类型、四分体的形态以及成熟花药壁的结构的差异, 揭示其在菊科植物系统分类与进化中的意义。
2018年全年和2019年4-10月, 在西北农林科技大学校园采集向日葵族向日葵(Helianthus annuus)、苍耳(Xanthium sibiricum)、金鸡菊(Coreopsis drummondii)、鬼针草(Bidens pilosa)、松果菊(Echinacea purpurea)、黄秋英(Cosmos sulphureus), 春黄菊族滨菊(Leucanthemum vulgare), 菜蓟族飞廉(Carduus nutans)、矢车菊(Centaurea cyanus), 堆心菊族天人菊(Gaillardia pulchella), 菊苣族蒲公英(Taraxacum mongolicum)、苦苣菜(Sonchus oleraceus)、黄鹌菜(Youngia japonica)、苦荬菜(Ixeris polycephala)。在宁陕县火地塘林场采集向日葵族牛膝菊(Galinsoga parviflora), 菊苣族毛连菜(Picris hieracioides), 菜蓟族牛蒡(Arctium lappa), 千里光族蒲儿根(Sinosenecio oldhamianus), 旋覆花族大花金挖耳(Carpesium macrocephalum)。采集不同大小的头状花序, 小的花序直接固定, 大的花序十字切割后固定。固定液选择卡诺氏固定液, 纯酒精:冰醋酸=3:1(V:V), 固定24 h后转入70%酒精中保存。制片前采用爱氏苏木精整体染色或者苯胺番红整体染色, 常规石蜡切片法制片, Motic摄影显微镜观察并拍照。
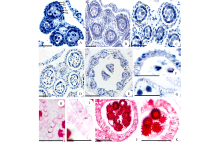
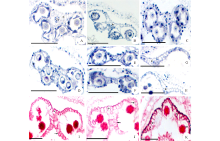
在花粉母细胞时期花药壁的结构相对较完整, 随着花粉母细胞发育成花粉粒, 药壁的结构会逐渐发生变化, 最常见的变化是绒毡层和中层细胞会逐渐解体, 药室内壁细胞纤维状加厚, 药室开裂。菊科植物花药横切后通常由4个花粉囊组成, 在花粉母细胞时期, 花粉囊壁由4层细胞组成, 从外向内依次为表皮、药室内壁、中层和绒毡层, 各层均由1层细胞组成(图1A~C)。中层细胞解体较早, 在花粉母细胞时期可以看到少量的中层细胞(图1A~C), 在四分体时期, 中层细胞已全部解体(图1D)。在菊科植物中, 也有由2个花粉囊组成的花药(图1E)。随着花粉粒的形成, 药室内壁细胞首先沿着径向壁伸长(图1F), 其次径向壁和内切向壁木质化增厚, 即“ U” 形增厚(图1H)。多数菊科植物在“ U” 形增厚的基础上, 又沿着径向壁方向呈条带状木质化增厚(图1I~K)。在舌状花亚科植物中, 药隔及其附近表皮下方的一层薄壁细胞也发生“ U” 形增厚, 不同的是径向壁和外切向壁发生木质化增厚, 呈倒“ U” 形增厚(图1G)。花药的表皮细胞在花粉粒发育过程中普遍解体消失(图1F~K)。
 | 图1 花药和药壁的结构 花药发育时期:A~C: 花粉母细胞时期; D: 四分体时期; E: 单核花粉粒时期; F~K: 成熟花粉粒时期。A:天人菊; B:黄秋英; C:苍耳; D:松果菊; E:黄鹌菜; F:松果菊; G:苦荬菜, * 示“ U” 形增厚; H:滨菊; I:矢车菊, * * 示条带状增厚; J:向日葵; K:金鸡菊。Ep:表皮; En:药室内壁; T:绒毡层; Ms:花粉母细胞; ML:中层; Co:药隔; Tds:四分体; Ls:花粉囊。 标尺=50 μ m。Fig.1 The structure of anther and anther wall Anther development period:A-C: meiocytes period; D: Tetrads period; E: Uninucleate pollen grains period; F-K: Mature pollen grains period. A: G. pulchella; B: C. sulphureus; C: X. sibiricum; D: E. purpurea; E: Y. japonica; F: E. purpurea; G: I. polycephala, * show“ U” thickening; H: L. vulgare; I: C. cyanus, * * show strip thickening; J: H. annuus; K: C. drummondii. Ep: Epidermis; En: Endothecium; T: Tapetum; Ms: Meiocytes; ML: Middle layer; Co: Connectivum; Tds: Tetrads; Ls: Locules. Scale bars=50 μ m. |
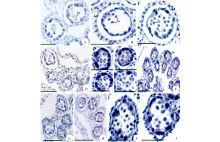
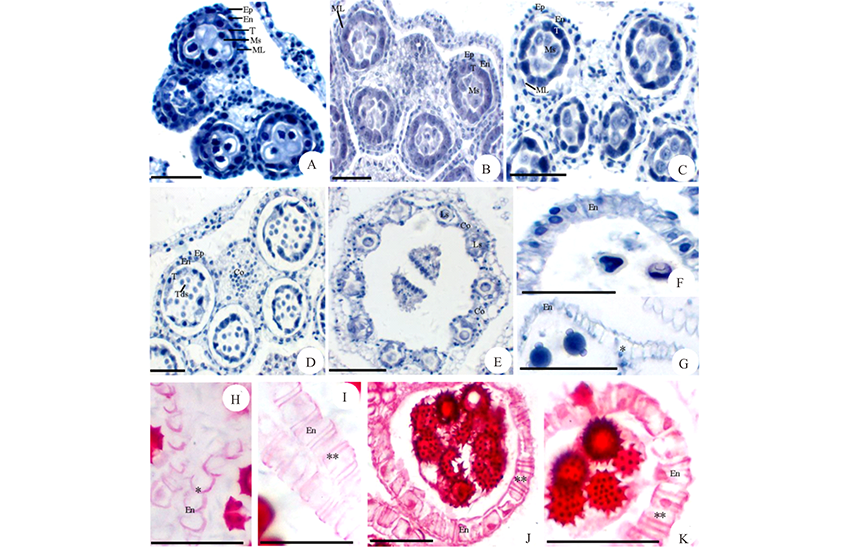
通过对菊科8属植物花粉母细胞减数分裂过程的观察, 发现菊科植物花粉母细胞胞质分裂类型有2种, 连续型和同时型。如牛蒡(图2A)、松果菊(图2B, C)和大花金挖耳(图2D)的花药, 花粉母细胞减数分裂的第一次分裂后产生分隔壁, 形成含两个子细胞的二分体, 第二次分裂后形成四分体, 花粉母细胞减数分裂胞质分裂类型属于连续型。在苍耳(图2E)的花药中, 可以看到花粉母细胞减数分裂的4个不同时期, 从细胞形态可以判断, 花粉母细胞减数分裂的胞质分裂类型属于同时型。从蒲儿根(图2F)花药中不同时期四分体的形态可以推断, 其花粉母细胞减数分裂的胞质分裂类型也属于同时型。
 | 图2 胞质分裂类型和四分体形态 花药发育时期:A~J: 二分体和四分体时期。A:牛蒡; B, C:松果菊; D:大花金挖耳; E:苍耳; F:蒲儿根; G:牛膝菊; H:毛连菜; I, J:天人菊。Tds:四分体; Dds:二分体。标尺=50 μ m。Fig.2 Cytokinesis type and tetrad morphology Anther development period: A-J: dyads and tetrads period. A: A. lappa; B, C: E. purpurea; D: C. macrocephalum; E: X. sibiricum; F: S. oldhamianus; G: G. parviflora; H: P. hieracioides; I, J: G. pulchella. Tds: Tetrads; Dds: Dyads. Scale bars=50 μ m. |
菊科植物小孢子四分体的形态以四面体形为主(图2G~J), 也发现有少数的十字形(图2G, J)。
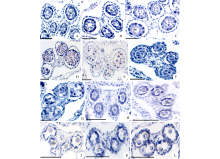
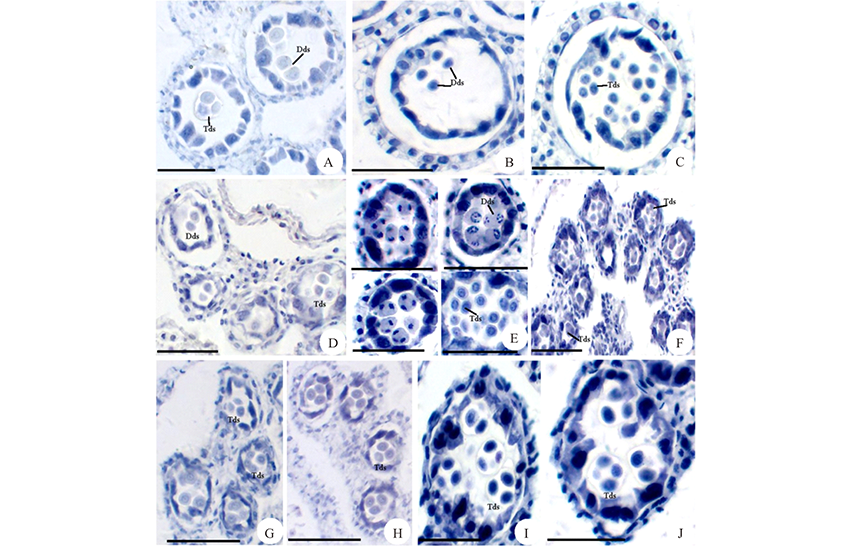
根据花药绒毡层细胞形态变化的观察, 菊科植物花药绒毡层有两种类型:变形绒毡层和分泌绒毡层。以苍耳为例, 说明变形绒毡层的形态变化过程。在花粉母细胞时期(图3A)和四分体时期(图3B), 绒毡层细胞整齐地排列在花粉囊壁的最内层, 围绕着花粉母细胞或者四分体。在四分体分离形成小孢子时期(图3C), 绒毡层细胞沿着径向壁伸长, 随着细胞壁的溶解, 细胞内的原生质进入花粉囊中, 充满单核花粉粒之间的空隙(图3D), 最后在花粉粒之间解体(图3E)。在天人菊花药中, 花粉母细胞时期绒毡层细胞结构完整, 排列在花粉母细胞外围(图3F), 在单核花粉粒发育到单核靠边期时, 绒毡层细胞才开始进入花粉囊(图3G), 发生的时间相对苍耳较晚, 也属于变形绒毡层。在毛连菜花药中, 花粉母细胞时期(图3H)和四分体时期(图3I)的绒毡层细胞含有较浓厚的原生质, 分布在花粉母细胞或者四分体的外围。在单核花粉粒时期(图3J), 绒毡层细胞仍位于原来的位置, 细胞中的原生质明显减少, 细胞正在解体, 直至解体消失。因此, 毛连菜的绒毡层属于分泌绒毡层。苦荬菜的绒毡层细胞, 从四分体时期(图3K)发育到单核花粉粒后期(图3L), 绒毡层细胞在原来的位置逐渐解体, 也属于分泌绒毡层。
 | 图3 花药绒毡层细胞的形态变化 花药发育时期:A, F, H:花粉母细胞时期; B, I, K:二分体和四分体时期; C, D, J:单核花粉粒时期; E, G, L:单核靠边期。A~E:苍耳; F~G:天人菊; H~J:毛连菜; K~L:苦荬菜。T:绒毡层; UPG:单核花粉粒; Ms:花粉母细胞; Tds:四分体。 标尺=50 μ m。Fig.3 The morphological changes of tapetum cells Anther development period: A, F, H: Meiocytes period; B, I, K: Dyads and tetrads period; C, D, J: Uninucleate pollen grains period; E, G, L: Single nucleus sideline period. A-E:X. sibiricum; F-G: G. pulchella; H-J: P. hieracioides; K-L: I. polycephala. T: Tapetum; UPG: Uninucleate pollen grains; Ms: Meiocytes; Tds: Tetrads. Scale bars=50 μ m. |
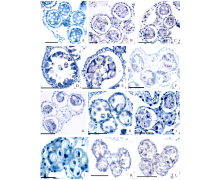
通过对管状花亚科向日葵族、菜蓟族、春黄菊族、堆心菊族和千里光族8种植物花药绒毡层形态的观察, 发现变形绒毡层是管状花亚科植物的共同特点(图4)。向日葵族是管状花亚科较原始的类型, 花药绒毡层细胞进入花粉囊的时间比较早, 发生在小孢子四分体分离形成小孢子的时期。绒毡层细胞的原生质进入花粉囊后融合形成周原质团, 包裹着单核花粉粒(图4A, B), 周原质团在花粉囊中存在的时间相对较长。在菜蓟族中, 绒毡层细胞进入花粉囊的时间相对较晚, 发生在单核花粉粒后期, 即单核靠边期(图4D, F), 绒毡层细胞的原生质进入花粉囊后也融合形成周原质团, 包裹着花粉粒(图4E, G)。在春黄菊族和堆心菊族中, 绒毡层细胞的原生质在单核靠边期时进入花粉囊中(图4H), 原生质发生部分融合, 松散地分布在花粉粒周围(图4I, J)。在千里光族中, 绒毡层细胞的原生质在单核靠边期时径向伸长进入花粉囊(图4K), 原生质很少发生融合, 只是分散在花粉粒之间, 最后解体消失(图4L)。
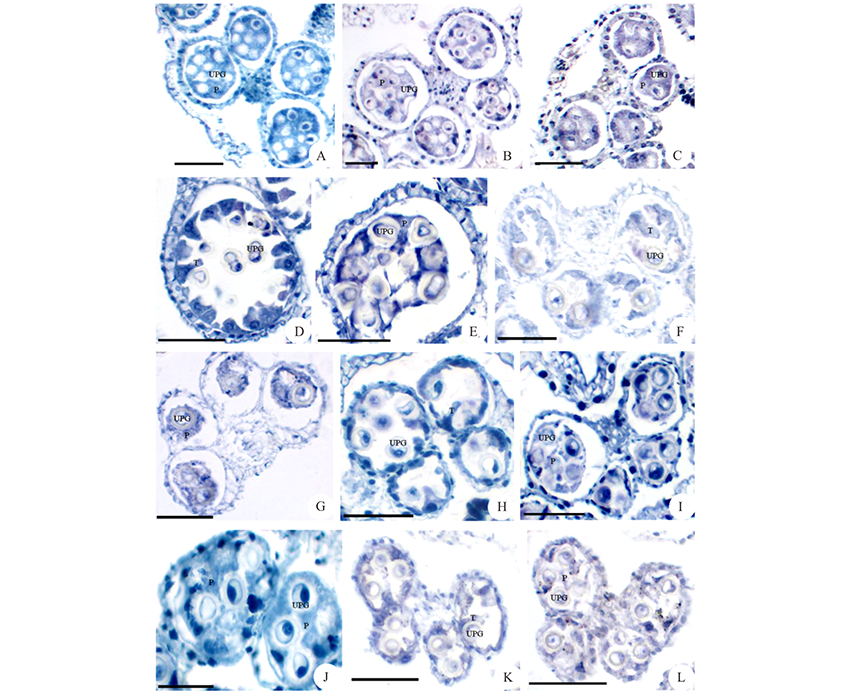 | 图4 管状花亚科植物的花药绒毡层类型 花药发育时期:A~C:单核花粉粒时期; D~L:单核靠边期。A~C:向日葵族(A:金鸡菊; B:松果菊; C:鬼针草); D~G:菜蓟族(D~E:矢车菊; F~G:飞廉); H~I:春黄菊族滨菊; J:堆心菊族天人菊; K~L:千里光族蒲儿根。UPG:单核花粉粒; P:周原质团; T:绒毡层。标尺=50 μ m。Fig.4 The tapetum type of Carduoideae Anther development period:A-C: Uninucleate pollen grains period; D-L: Single nucleus sideline period. A-C: Heliantheae (A: C. drummondii; B: E. purpurea; C: B. pilosa); D-G: Cynareae(D~E: C. cyanus; F~G: C. nutans); H~I: L. vulgare of Anthemideae; J:G. pulchella of Helenieae; K~L:S. oldhamianus of Senecioneae. UPG: Uninucleate pollen grains; P: Periplasmodium; T: Tapetum. Scale bars=50 μ m. |
通过对舌状花亚科菊苣族5个属5种植物毛连菜、蒲公英、苦苣菜、苦荬菜和黄鹌菜花药结构的观察, 发现舌状花亚科植物花药绒毡层的类型普遍属于分泌绒毡层, 绒毡层细胞在原来的位置解体直至完全消失(图5A~F)。当花粉粒成熟以后, 药室内壁细胞内切向壁和径向壁纤维加厚(图5I, J, 箭头所示), 与此同时, 药隔外侧表皮下方一层细胞也纤维加厚(图5G~K, * 所示), 与药室内壁形成的纤维细胞层相连接, 有利于药室从内侧打开。
 | 图5 舌状花亚科植物的花药绒毡层类型和药隔的结构 花药发育时期:A~F:单核花粉粒时期; G~K:成熟花粉粒时期。A:毛连菜; B, I:蒲公英; C, G:苦苣菜; D, H, J:苦荬菜; E, F, K:黄鹌菜。T:绒毡层; Co:药隔。“ * ” 示药隔外侧表皮下方细胞纤维化增厚。“ → ” 示药室内壁细胞纤维化增厚。标尺=50 μ m。Fig.5 The tapetum type and connectivum structure of Cichorioideae Anther development period: A-F: Uninucleate pollen grains period; G-K: Mature pollen grains period. A: P. hieracioides; B, I: T. mongolicum; C, G: S. oleraceus; D, H, J: I. polycephala; E, F, K: Y. japonica. T: Tapetum; Co: Connectivum.“ * ” shows that the cells below the epidermis of connectivum were fibrotic and thickened.“ → ” shows that the endothecium were fibrotic and thickened. Scale bars=50 μ m. |
根据绒毡层发育后期的形态变化, 将绒毡层分为两种类型:分泌绒毡层和变形绒毡层[20], 本研究表明, 向日葵族植物花药绒毡层细胞在小孢子形成时就进入花粉囊, 并在花粉囊中融合形成周原质团, 包裹着小孢子, 随着小孢子发育成花粉粒, 周原质团逐渐解体消失。由于周原质团在花粉囊中存在的时间相对较长, 通过切片很容易观察到, 因此, 绒毡层的类型也容易判别, 属于变形绒毡层, 与前人的研究结果也比较一致。据研究报道, 向日葵族植物牛膝菊、三裂叶豚草和豚草的花药绒毡层均属于变形绒毡层[13, 14, 16]。在菜蓟族、春黄菊族和堆心菊族中, 绒毡层细胞进入花粉囊的时间相对较晚, 发生在单核花粉粒后期, 即单核靠边期, 绒毡层细胞的原生质进入花粉囊后, 菜蓟族植物原生质全部融合形成周原质团, 而春黄菊族和堆心菊族植物原生质部分发生融合形成周原质团。因此, 只要观察时间准确, 也很容易判断绒毡层的类型。据研究报道, 堆心菊族黄顶菊和孔雀草的绒毡层类型为变形绒毡层[11, 12]。在千里光族中, 绒毡层细胞在单核靠边期时才径向伸长进入花粉囊, 原生质很少发生融合, 只是分散在花粉粒之间, 最后解体消失, 绒毡层的类型是变形绒毡层向分泌绒毡层的过渡状态, 兼有变形绒毡层和分泌绒毡层的形态。因此, 关于千里光族植物绒毡层类型的研究报道结果不一致。据研究报道, 千里光族植物款冬的绒毡层细胞在单核花粉粒发育后期才突入药室腔, 绒毡层细胞不发生融合, 属于分泌绒毡层[6]。但从本研究对绒毡层细胞形态变化的描述, 其应该属于变形绒毡层。据报道, 千里光族千里光的花药具有4个花粉囊, 绒毡层为变形绒毡层[15]。紫菀族、金盏花族和旋复花族植物花药绒毡层的发育形态与千里光族很相似, 研究结果不一致。据研究报道, 紫菀族小蓬草的花药具有分泌绒毡层[8], 也有报道紫菀族植物是变形绒毡层和分泌绒毡层共存[17]。金盏花族金盏花花药绒毡层主要是变形绒毡层, 还有少量的分泌绒毡层[18]。旋复花族革苞菊的花药绒毡层属于变形绒毡层[10]。研究结果不一致的主要原因是紫菀族、金盏花族和旋复花族植物花药绒毡层细胞在单核花粉粒后期才进入花粉囊。从小孢子形成至单核花粉粒后期, 绒毡层细胞在原来位置慢慢解体; 从单核花粉粒后期至花粉粒成熟, 绒毡层细胞进入花粉囊中完全解体, 兼有分泌绒毡层和变形绒毡层两种特性。另外, 绒毡层细胞进入花粉囊后不发生融合也是影响绒毡层发育类型判定的原因。
菊苣族属于舌状花亚科。本研究对菊苣族5个属5种植物毛连菜、蒲公英、苦苣菜、苦荬菜和黄鹌菜花药结构发育动态进行观察, 根据绒毡层细胞的形态变化, 确定菊苣族植物的花药绒毡层类型为分泌绒毡层。据研究报道, 菊苣族台湾翅果菊的绒毡层类型为分泌绒毡层[7], 而菊苣菜的绒毡层类型为变形绒毡层[19], 本研究结果与此不完全一致。研究结果不一致的主要原因有两点:一是菊苣族植物花药细长, 花药花粉囊腔很小, 常常见到一个花粉囊中只有1粒花粉粒, 绒毡层细胞紧贴着花粉粒, 很难判断绒毡层的发育类型。在选择试验材料时, 选取花粉囊腔较大的材料, 可以解决这个问题。二是观察时期较少, 只有通过动态观察绒毡层细胞形态的变化, 才能得出准确的结论。
根据菊科植物的系统关系分支图[21, 22], 向日葵族是菊科植物中较原始的类群, 菊科各族的演化趋势为:泽兰族→ 向日葵族→ 堆心菊族→ 千里光族→ 紫菀族→ 金盏花族→ 旋复花族→ 菊苣族。在演化趋势中前7个族属于管状花亚科, 而菊苣族属于舌状花亚科, 因此, 可以推断舌状花亚科较管状花亚科进化。通过本研究和前人对向日葵族、堆心菊族、千里光族、紫菀族和金盏花族花药绒毡层发育过程的观察, 发现菊科植物从变形绒毡层、经过变形绒毡层向分泌绒毡层的过渡类型逐渐演化成菊苣族的分泌绒毡层。向日葵族植物具有典型的变形绒毡层, 绒毡层细胞从小孢子时期进入花粉囊, 并融合形成周原质团包围着花粉粒。千里光族和旋覆花族植物从单核花粉粒后期开始绒毡层细胞才进入花粉囊, 且细胞不发生融合, 属于过渡类型。菊苣族植物花药绒毡层细胞原位解体, 属于分泌绒毡层。研究表明, 变形绒毡层是菊科管状花亚科的绒毡层类型, 而分泌绒毡层是菊科舌状花亚科的绒毡层类型。变形绒毡层的形态在管状花亚科不同族之间存在明显差异, 绒毡层的发育类型和形态可以作为菊科亚科和族分类的依据之一。
根据花冠类型和植物体是否含有乳汁, 菊科分为两个亚科:管状花亚科和舌状花亚科。本研究表明, 菊科植物花药普遍由4个花粉囊组成, 也发现有2个花粉囊组成的花药。花粉囊壁由表皮、药室内壁、中层和绒毡层4层细胞组成, 中层细胞解体较早。随着花粉粒的形成, 药室内壁先“ U” 形增厚, 再条带状增厚。花粉母细胞胞质分裂类型有连续型和同时型2种, 小孢子四分体以四面体形为主。绒毡层的发育类型有变形绒毡层和分泌绒毡层两种类型, 变形绒毡层是管状花亚科植物的共同特点, 分泌绒毡层是舌状花亚科植物的共同特点。从向日葵族向菊苣族的逐渐演化过程中, 绒毡层的发育类型从向日葵族的变形绒毡层、经过变形绒毡层向分泌绒毡层的过渡类型逐渐演化为菊苣族的分泌绒毡层。变形绒毡层的形态在管状花亚科不同族之间存在明显差异, 绒毡层的发育类型和形态可以作为菊科亚科和族分类的依据之一。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|