


{kind=link}
{kind=link}
{kind=link}
{kind=link}
借助抽雄期长茎段育苗释放多年生饲草玉米茎节扩繁潜力
[严旭1, 2, 3  , 吴子周
, 吴子周1 , 李影正1 , 李晓锋1 , 何如钰1 , 杨春燕1, 4 , 李杨1 , 周阳1 , 石建忠1 , EBENEZER Kofi Sam1 , 何建美1 , 荣廷昭1 , 唐祈林1, * ]
, 吴子周]
|
|


作者简介:严旭(1987-),男,四川仪陇人,博士。E-mail: pratum@qq.com
多年生饲草玉米‘玉草6号’为分蘖多、生物量高、适口性好的C4丛生型高大禾草,仅能通过营养体无性繁殖。越冬返青幼苗(分株苗)和上年茎节成苗(茎节苗)是其潜在繁殖体,但其茎节苗的田间表现和茎节田间育苗潜能未知。田间建植分株苗和茎节苗,调查繁殖体类型对植株形态和生物量的影响;田间播种茎节,调查茎节播期、位置和茎段大小对茎节育苗的影响。茎节苗建植成功率(97.5%)高于分株苗(87.5%),两种繁殖体类型均可安全越冬(越冬率100%)。建植当年,分蘖期茎节苗植株茎径显著高于分株苗;此外,茎节苗植株的株高、草长、茎径、基部周长和分蘖均有微弱优势,但繁殖体类型间无显著差异;生物量在繁殖体类型间也无显著差异。次年,植株形态、生物量在繁殖体类型间也无显著差异。茎叶比年际间无显著差异,茎径次年显著降低,其他形态和产量性状均显著高于建植第1年。茎节播期和位置对茎节成活率、扩繁系数和种苗大小有极显著影响,而茎段大小对上述育苗参数无显著影响。在各组合中,抽雄期中、下部长茎段育苗表现最佳。茎节繁殖能拓宽多年生饲草玉米的繁殖体类型,并能提高建植成功率且不影响饲草生产性能。采用抽雄期中、下部长茎段育苗是释放多年生饲草玉米茎节扩繁潜力的有效措施。
, WU Zi-zhou
Novel perennial forage maize cultivar (‘Yucao No.6’, Yu6), an allopolyploid containing genomes of Zea mays, Tripsacum dactyloides and Zea perennis, is a palatable, sterile, high-tillering and productive C4 grass. By virtue of sterility, it is limited to vegetative propagation. In comparison with ramet division as a propagation method, little is known about the performance of plants propagated from stem segments and the clonal potential of stem. The aim of this study was to clarify the field performance of plants derived from stem segments and the ability of stem to produce new plants. Field trials were performed to investigate the effect of propagule type on the developmental morphology and above-ground biomass (AGB) of Yu6 over two years, and to evaluate the effects of sowing time, nodal position and stem segment size on the regeneration of aboveground stem fragments from Yu6 in a field setting. Results showed that the establishment mortality (12.5%) of ramet division was higher than that (2.5%) of stem division in the establishment year, and that both types of propagules could safely overwinter (100% overwintering rate in the first winter after planting). We found slight advantages in plant height (PH), grass length (GL), basal circumference (BC), tiller number (TN), AGB for stem propagation compared to ramet division, and no propagule-related difference in stem to leaf ratio (SLR) in the establishment year, except that stem diameter (SD) was significantly greater in stem propagated plants during the tillering stage. There was no significant difference in plant characteristics (PH, GL, BC, TN, SD and SLR) and AGB between the propagule types in the second year following planting, but the second-year metrics (except SLR and SD) were significantly higher than the first year. Sowing date and nodal position had a significant effect on the survival rate, multiplication rate and plantlet size. However, the effect of stem segment size on these parameters was non-significant. Of the treatment combinations, the longer stem segment of the basal and middle stems at the tasseling stage showed the best plantlet regeneration performance. Briefly, use of stem propagation can enhance the propagation opportunities for perennial forage maize, and increase the success rate of establishment without compromising forage production performance. The most effective propagation methodology was the raising of plantlets from the longer stem segments of perennial forage maize at the tasseling stage. Adoption of this methodology could increase the popularity and use of perennial forage maize.
新型多年生饲草玉米(如‘ 玉草5号’ 和‘ 玉草6号’ )由玉米(Zea mays)、摩擦禾(Tripsacum dactyloides)与四倍体多年生大刍草(Zea perennis)经远缘杂交和染色体重组人工合成, 因其为杂合体, 后代性状变异大, 为了固定杂种优势, 采用无性繁殖[1, 2]。多年生饲草玉米少有根茎, 越冬器官为地下部茎秆(非根茎); 返青时地下茎秆根颈芽萌发, 植株再生。采挖、拆分和重植越冬植株是其当前主要扩繁途径(分株扩繁)。茎节扩繁具有提升多年生饲草玉米扩繁系数的潜力, 但其茎节扩繁植株的田间表现和茎节的田间扩繁潜能并不清楚。
繁殖体(种子、根茎、茎节或组培苗等)是禾本科作物实现田间建植与世代更迭的基础, 影响田间管理和作物生长、发育, 最终可能改变作物的产量与品质。如, 芦竹(Arundo donax)根茎和茎节扩繁影响建植当年植株茎高、茎径、茎秆密度和地上生物量[3]。芦竹腋芽快繁植株建植当年表现更多分蘖, 而腋芽成苗植株在株高和产量上显现优势[4]。虉草(Phalaris arundinacea)根茎繁殖在第2年即可获得饲草和种子高产, 种子繁殖则在第3年[5]。又如芒(Miscanthus spp.), ‘ 奇岗’ (Miscanthus× giganteus)建植期(1~3年)根茎扩繁植株基部周长更大, 而茎节扩繁植株分蘖更多[6]; 根茎发育植株较组培苗植株分蘖更少、茎秆和根茎更粗壮[4, 7, 8], 体细胞胚胎发生微繁苗定植后植株生长速度快于腋芽诱导的微繁苗[8]; 同时根茎扩繁具有更高的建植成功率和越冬率[9]。上述芒不同繁殖体建植后植株表现出的形态差异并未影响最终产量[4, 6, 7, 8, 9]; 但Lewandowski[8]发现根茎扩繁植株根茎中氮、磷、钾和糖含量更高, 而微繁植株地上部的氮、钾、水分和灰分含量更高。在Pł azek等[10]的研究中, 奇岗根茎扩繁植株的茎秆高度、干物质积累更高; 而组培苗建成植株的分蘖、半纤维素和灰分含量更高。
茎节源自禾草地上部, 数量巨大、无须采挖、环境友好, 相较于分株其潜在的扩繁系数更高, 被视为更具发展潜力的繁殖体[3]。对10份柳枝稷(Panicum virgatum)茎节萌芽的研究发现[11], 各种质下部茎节的出芽率最高(60%~98%); 其中, 3份种质的中、上部茎节萌芽率分别超过80%和50%, 7份种质中、上部茎节萌芽率分别小于40%和10%。玉草5号上、中和下部单个茎节扦插50 d后成活率依次为10.32%、26.93%和51.69%[1]。‘ 玉草1号’ (Z. mays× Z. perennis)含2个节的茎段生根率和成活率均高于单个茎节[12]。奇岗茎节出芽和生根能力与植株的生长年限无关, 但与茎节收获期密切相关[13]; 其下部茎节的生根率较上部茎节更高, 单个茎节插条扩繁系数更高, 而包含2个茎节的茎段其幼苗根系发育更好[14]。芒草茎节春、夏季均可出芽、生根, 茎节成苗率春季最高(34%), 其次为仲夏(29%), 夏末最低(25%), 秋、冬季茎秆衰老使其丧失了出芽和生根能力; 而芦竹(Arundo donax)茎节全年具有出芽和生根能力[15]。由此可见, 禾草茎节扩繁受物种、基因型、茎节位置、出芽率、生根率、茎段大小和发育阶段等多种因素的影响。
茎节扩繁能提升芦竹[3, 4, 15, 16]、芒[3, 4, 15, 16, 17]和柳枝稷[11]等禾草的扩繁系数。茎节可作为多年生饲草玉米[1, 12]及其亲本四倍体多年生大刍草[18]的繁殖体也被证实, 但茎节作为繁殖体如何影响多年生饲草玉米的田间表现未知, 同时对其茎节田间育苗效果知之甚少。本研究旨在明确:1)茎节苗、分株苗两类繁殖体对多年生饲草玉米建植成功率和越冬率、形态发育以及生物量的影响; 2)茎节育苗时间、茎节位置及茎段大小对茎节田间育苗的影响, 以期为多年生饲草玉米茎节扩繁的利用提供信息。
试验地位于四川农业大学温江基地(30° 71' N, 103° 87' E, 海拔536 m), 水稻土, pH趋于中性至弱碱性。耕作层有机质含量2.29%、全氮0.17%、全磷0.056%和全钾2.38%。试验期间降水量与温度数据从中国气象数据网(http://data.cma.cn/)获取并整理。降水和高温集中在6-8月, 雨热同期(图1)。
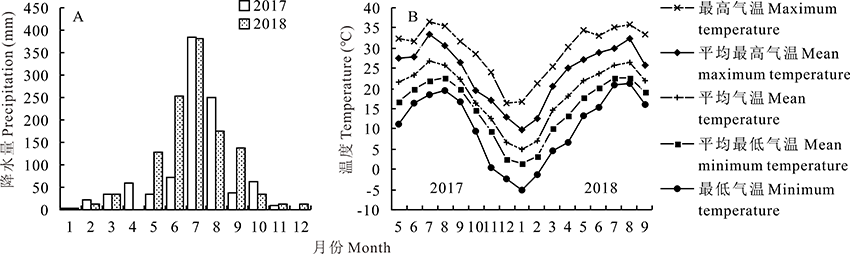 | 图1 试验期间试验地降水量和温度Fig.1 Precipitation and temperature data during the trial in research site |
试验采用完全随机区组设计, 4次重复。精细整地, 小区面积4 m× 6 m, 小区间隔1.5 m, 株行距1.0 m× 1.2 m, 各小区20株。2017年5月9日, 取形态相似、株高约10 cm的玉草6号茎节苗(取自2016年茎节育苗再生植株)和分株苗(拆分自越冬植株的返青幼苗), 剪去约2/3叶片后定植, 小区外定植相应繁殖体。定植深度约5 cm, 覆土后浇水, 新叶出现前常浇水以应对移栽冲击和组织失水。定植后45 d调查建植成功率(小区同类型繁殖体存活植株数/80), 并将小区外植株带土移栽以替换区内死亡植株, 并移除区外植株。定植后60 d小区内随机选定3株挂牌用于形态测定, 之后15 d测定一次, 直至抽雄期测产。2018年返青后测越冬率(小区同类型繁殖体存活植株数/80), 在抽雄期测定植株形态和产量性状。每年穴施200 kg· hm-2尿素, 拔节期施1/3, 大喇叭口期施2/3。其他管理同大田生产。
形态性状包括分蘖数(单株茎秆数)、株高(主茎端部叶颈或花序顶端沿茎到茎基部的距离, cm)、草长(主茎端所有叶沿茎端拉直, 茎基部沿茎到最长叶尖的距离, 当该值小于株高时草长记为株高, cm)、茎径(主茎地上第1节节间中部的长轴和短轴的平均值, mm)和基部周长(皮尺紧贴地面围绕单株所有分蘖测得的长度, cm), 按小区平均后用于后续分析。测产时, 小区内部植株刈割称鲜重, 再随机取约1 kg鲜样称重后65 ℃烘干至恒重。烘干样分离茎(茎秆、叶鞘和雄花序)、叶(叶片)分别称重, 茎叶比=茎干重/叶干重。根据刈割面积、鲜重和干物质含量折算鲜、干草产量。
以2018年1.0 m× 1.2 m株行距下建植的玉草6号地上部茎秆为扩繁材料。8月15日起每隔约30 d留茬5 cm收获茎秆, 直至11月截止。剔除叶片、侧枝(9月底出现)和茎秆端部4节后, 按茎秆茎节数(n)划分茎节位置。下部茎节(basal stem, B)为茎秆基部1~[n/3](向下取整)节, 上部茎节(apical stem, A)为茎秆端部1~[n/3](向下取整)节, 剩余为中部茎节(middle stem, M)。区分的各位置茎节即为“ 长茎段” , 每段包含2~5个茎节(long segment with 2-5 nodes, L); 茎段再从各节间中部剪伐后即成“ 单茎节” (short segment with one node, S)。试验包括4个时期(8、9、10和11月)、2种茎段大小(L和S)和3种茎节位置(B、M和A), 共计24个处理, 每个处理3次重复, 每个重复包含50个茎节(即, L为包含50个茎节的长茎段, S为50个单茎节)。
茎秆收获和育苗在2 d内完成。育苗于温江试验基地(同上)进行, 犁、耙至土壤疏松、平整。设置3个平行小区, 间隔1 m, 小区面积2 m× 24 m。按行距20 cm开沟, 沟深约5 cm。各小区内设置1个重复, 处理间随机排列, 同一处理茎节于连续5行中均匀放置, 覆土后使用二甲戊灵(1237.5 g· hm-2)表土喷雾抑制杂草萌发。最后一次(11月)育苗时, 各小区人工除草后覆PVC (polyvinyl chloride)透明薄膜, 膜厚0.02 mm, 脊高1 m。将热电偶温度计(宇问SSN-61, 深圳)放入拱棚内中心点, 设置30 min记录1次温度, 期间膜内最低气温为0.6 ℃(2019年1月16日)。于3月25日, 调查记录地面可见幼苗。茎节成活率=(成苗的茎节个数/50)× 100%, 茎节扩繁系数=可移栽的种苗个数/50, 即单个茎节能生成的种苗数。种苗大小以株高表示, 每个重复内随机取3株以平均值表示用于后续分析; 重复内再生茎节小于3时, 测2或1株。
在R 3.4.3(R development core team, 2017)中采用agricolae分析数据, 用Excel 16.0绘图。采用卡方检验(Chi-square test)比较繁殖体类型间的建植成功率和越冬率差异; 采用t检验比较繁殖体类型间的形态差异; 采用方差分析(ANOVA)比较繁殖体类型和生长年限对多年生饲草玉米植株形态和生物量的影响; 采用非参数检验(Kruskal-Wallis)比较育苗时间、茎节位置及茎段大小对茎节育苗的影响, 图表中的数据以平均值± 标准误表示。
2.1.1 幸存能力 茎节扩繁建植成功率(97.5%)高于分株扩繁(87.5%)(P=0.4454)。分株和茎节扩繁植株越冬率均为100%, 越冬期间最低气温为-5.1 ℃(图1B), 表明多年生饲草玉米越冬性能不受繁殖体类型影响且其具有一定的抗寒性。
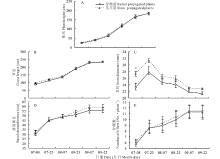
2.1.2 形态发育 多年生饲草玉米建植第1年不同形态性状的发育策略并不一致(图2)。株高和草长在分蘖期增长缓慢, 在拔节期迅速增加(图2A, B)。茎径、基部周长和分蘖数在分蘖期生长最为迅速, 在拔节期分蘖数、基部周长增速减缓, 而茎径在拔节期后逐渐缩小(图2C~E)。进入生殖阶段直至抽雄期植株形态性状的增长几乎停滞(图2)。定植后60 d茎节苗茎径显著高于分株苗, 这一移栽初期茎径表现出的差异贯穿整个生育期, 但差异逐渐减小(图2C); 其他形态性状在各测定时期繁殖体间无显著差异, 但茎节苗有微弱优势(图2)。次年, 除茎径显著降低外, 其他形态性状均极显著高于建植第1年; 刈割时繁殖体类型对各形态性状无显著影响(表1)。
| 表1 繁殖体类型对多年生饲草玉米抽雄期形态特征的影响 Table 1 Effects of propagule type on morphological characteristics of the perennial forage maize at the tasseling stage |
2.1.3 产量性状 分株扩繁和茎节扩繁的干草产量建植第1年分别为10921和11402 kg· hm-2, 次年分别为19755和19793 kg· hm-2(表2)。由此可见, 同一年份繁殖体类型间干草产量无显著差异, 建植第2年干草产量极显著提升。鲜草产量与干草产量的变化规律相似, 茎叶比未受繁殖体类型和年份的影响(表2)。结果表明多年生饲草玉米产量性状不受繁殖体类型影响。
| 表2 繁殖体类型对多年生饲草玉米产量性状的影响 Table 2 Effects of propagule type on yield traits of the perennial forage maize |
2.2.1 组合效应 田间育苗条件下, 各处理组合对茎节育苗各参数有极显著影响(P< 0.0001)(表3)。茎节成活率最佳处理组合集中在9月, 该月最佳处理组合占所有最佳处理组合的60%。茎节发育形成的种苗基部可进一步分蘖生成可移栽种苗(图3)。因此, 在09BL、09ML和09MS茎节成活率不足45%的情况下, 其茎节扩繁系数超过1。茎节扩繁系数最佳处理组合集中在9月, 该月最佳处理组合占所有最佳处理组合的67%。结果表明茎节播期对茎节扩繁系数有重要影响, 且9月茎节育苗效果最佳。种苗大小未受茎节位置和茎段大小的影响, 育苗时间对种苗大小有极显著影响, 8月的茎秆育苗培育的种苗最大。
| 表3 育苗时期、茎节位置和茎段大小的处理组合对茎节育苗的影响 Table 3 Effects of combination of sowing time, nodal position and stem segment size on plantlet formation |
 | 图3 抽雄期单茎节成苗(A)和其拆分的可移栽种苗(B)Fig.3 A plantlet developed from a single node at the tasseling stage (A) and five transplantable propagules derived from this plantlet (B) |
2.2.2 育苗时间 忽略茎节位置和茎段大小, 育苗时间对茎节成活率、茎节扩繁系数和种苗大小有极显著影响(P< 0.0001)(图4A~C)。茎节成活率9月最高, 平均为33.67%, 其后依次为10月(15.56%)、8月(14.44%)和11月(7.00%)。茎节扩繁系数9月最高, 平均为1.05, 其后依次为8月(0.57)、10月(0.33)和11月(0.08)。种苗大小随着育苗时间的推迟逐渐变小, 8月最高为23.39 cm, 其后依次为9月(11.81 cm)、10月(7.27 cm)和11月(4.37 cm), 彼此间差异显著。
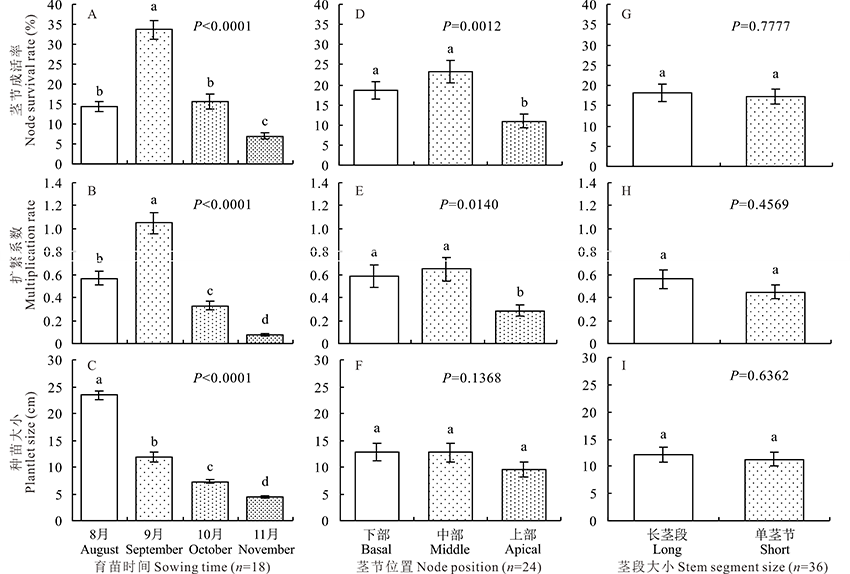 | 图4 育苗时间、茎节位置或茎段大小对茎节育苗的影响 不同小写字母表示该性状差异显著(P< 0.05)。Fig.4 Effects of sowing time, nodal position or stem segment size on plantlet formation Different lowercase letters indicate that the trait analyzed is significantly different at P< 0.05 level. |
2.2.3 茎节位置 忽略育苗时间和茎段大小, 茎节位置对茎节成活率(P=0.0012)、茎节扩繁系数(P=0.0140)和种苗大小(P=0.1368)的影响不同(图4D~F)。中、下部茎节成活率分别为23.33%和18.67%, 极显著高于上部茎节(11.00%)。中、下部茎节扩繁系数分别为0.65和0.59, 显著高于上部茎节(0.28)。中、下部茎节种苗大小分别为12.72和12.81 cm, 略高于上部茎节(9.60 cm)。
2.2.4 茎段大小 忽略育苗时间和茎节位置, 茎段大小对茎节成活率(P=0.7777)、茎节扩繁系数(P=0.4569)和种苗大小(P=0.6362)的影响均不显著(图4G~I)。长茎段和单茎节的茎节成活率、茎节扩繁系数和种苗大小分别为18.11%和17.22%、0.56和0.45以及12.12和11.30 cm。
建植成功率影响定植时建植密度和成本, 并影响收获时群体密度和产量[9]。多年生饲草玉米返青幼苗生长密集, 其根系和地上部长势均弱于茎节苗; 返青幼苗的采挖和拆分对根系的机械损伤严重, 使得分株建植成功率低。Pyter等[19]观测芒根茎扩繁建植损失率为12%, 与多年生饲草玉米分株建植损失率相当。Boersma等[9]发现奇岗茎节和根茎繁殖建植损失率在各地区间变幅为16.6%~33.8%, 两种扩繁方法建植损失率无显著差异, 而环境(建植地)对根茎扩繁建植成功率产生显著影响。可见, 禾本科无性扩繁作物的建植损失普遍存在, 提高建植成功率需要提升繁殖体的质量以及营造适宜繁殖体的移栽环境。
植株越冬期间极端低温为-5.1 ℃(图1B), 表明多年生饲草玉米具有一定的耐寒能力。覆膜条件下茎节再生植株也表现出一定的低温耐受能力(0.6 ℃)。四倍体多年生大刍草耐寒[20], 将摩擦禾引入玉米可提升幼苗的耐寒能力[21], 表明多年生饲草玉米继承了多年生亲本的抗寒性。由于越冬器官被土壤、植物凋落物覆盖, 气温并不能真实反映其耐寒性[22]。本试验在抽雄期刈割后再生植株遭遇冷害形成的枯草层和土壤的保温作用确保了植株安全越冬。因此, 多年生饲草玉米越冬器官的耐寒能力有待确定, 以为其种植范围扩展提供参考。
虽遗传构成相同, 但不同器官来源的繁殖体所处的生理、生化状态不同, 由此可能产生表型变异[17]。本试验多年生饲草玉米建植时分株苗和茎节苗大小相似, 但茎节苗发育植株的茎径高于分株苗。繁殖体(根茎和微繁苗)发育植株的茎径差异在芒属植物也曾被观测到, 被认为是繁殖体内残存的激素差异所引起的[4]。建植当年, 茎径先增加后逐渐缩小, 这一现象也存在于芒属植物中[6]。生活中不难发现, 甘蔗(Saccharum officinarum)茎秆基部茎径较中部细小。多年生饲草玉米基部茎秆也随着生长的纵向拉伸和包被叶鞘的脱落(测量茎径时包含包裹茎秆的叶鞘), 导致后期茎径逐渐缩小。次年, 分蘖的极显著增加导致了茎径降低。除茎径外, 其他形态性状在繁殖体类型间无显著差异, 但建植当年茎节苗发育进程中多数形态性状较分株苗有微弱优势(图2)。
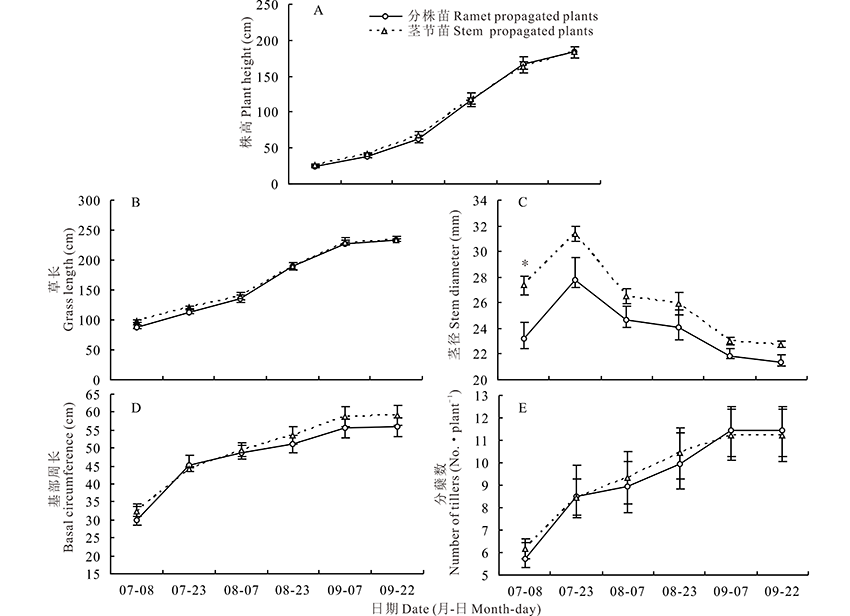 | 图2 建植第1年茎节苗和分株苗形态特征发育动态 “ * ” 表示该性状在繁殖体类型间差异显著(P< 0.05)。Fig.2 Dynamics of morphological characteristic of the perennial forage maize grown from stem propagatedplants and ramet propagated plants in the first growing season “ * ” indicates that the trait analyzed is significant difference at P< 0.05 level between propagule types. |
刈割时繁殖体间形态性状的一致导致繁殖体类型间的产量性状无显著差异, 但第2年产量性状极显著提升。建植当年产量较低的可能原因在于:1)移栽冲击和苗期干旱; 2)营养生长时间短, 受短日照诱导8月末植株开始生殖生长; 3)水分胁迫, 7月11日试验地遭遇强降水(温江气象站点记录该日降水量为108.5 mm), 植株根部受水分胁迫约1周。水分胁迫降低了生产性能, 但促使多年生饲草玉米生成的气生根确保了植株成活。试验地普通籽粒玉米全部死亡, 表明多年生饲草玉米遗传了亲本摩擦禾的耐涝能力[23]。次年产量极显著提升受惠于更长的营养生长期和适宜的生长环境(图1)。玉草5号(玉草6号姊妹系, 又名‘ 玉淇淋58’ )在温江2012年同期双株、高密度(1.0 m× 1.0 m)建植[1], 建植当年干草产量为22083 kg· hm-2, 表明提高密度和繁殖体数量可能提升种植当年饲草产量。
研究扩繁技术是无性系作物大规模应用的前提, 却常常被忽视。本试验在9月中旬(茎节处于抽雄期)育苗, 适宜的茎节成熟度(相较于8月)、温度和土壤墒情(相较于10-11月)共同促使该时期茎节育苗效果达到最佳。茎节出苗能力受茎节位置影响, 这似乎在禾草中普遍存在。如矮象草(Pennisetum purpurem)[24]和狼尾草(Pennisetum setaceum)[25]基部成苗率最高, 芦竹基部茎节的生根能力最强[3]。在多年生饲草玉米中, 上部茎节扩繁系数显著降低, 一种可能的解释是上部茎节中残留的激素倾向于诱导茎节分枝而非生根。中、下部茎节产生的种苗更大, 这可能与中、下部茎节中储备的营养物质含量更高有关。多年生饲草玉米并非每个茎节均能成苗, 在长茎段中未成苗茎节可能供给相邻已成苗茎节营养, 导致长茎段的育苗效果略好于单茎节。试验未发现长茎段会损失茎节扩繁系数, 同时长茎段能节省制作单个茎节的劳动力和时间成本。特别地, 多年生饲草玉米越冬时地上部遇霜死亡, 通过地下部茎秆根茎芽越冬。离体茎节形成的植株能否自然越冬, 次年再生成苗尚待验证。但在覆膜条件下茎节形成幼苗安全越冬有利于提早建植; 若推迟茎节苗移栽时间, 其形成的分蘖持续增多, 有可能进一步提升茎节扩繁系数。
茎节扩繁以地上茎节为繁殖体, 避免了分株对土壤和建成植株的破坏; 茎秆的收获、茎节苗的采挖以及茎秆和茎节苗的运输也使得繁殖体的传播更加方便、快捷。多年生饲草玉米返青植株拆分成单株后植株根系受损严重, 直播时1个越冬植株通常拆分成5~10株以确保种苗成活。这导致分株扩繁系数(5~10)低。玉草6号建植当年(1.0 m× 1.2 m)单株茎节数约224节, 在育苗效果最佳的9月育苗(茎节扩繁系数=1.05), 则单株茎节的扩繁系数约为235。相较于分株, 单株茎节的扩繁系数提升约20~50倍。芒等同类型无性系作物的微繁苗价格高昂[26]; 且有研究显示微繁苗冬季死亡率高, 并会降低建植期植株生产性能[8]。综上, 茎节无疑是当前多年生饲草玉米大规模推广的首选繁殖体。
茎节田间育苗受茎节发育时期、土壤水分和温度等多重因素影响。结合当前生产需求, 本试验仅关注3种重要因素对茎节田间育苗的影响, 其差异形成的生理、生化等机制有待研究。如赤霉素(gibberellic acid)可促进竹(Bambusa vulgaris)芽的增殖, 而细胞分裂素(cytokinin)则促进竹芽萌发[27]。生长素(4-indol-3-ylacetic acid)可提升芒地上部茎节芽和根的发生[24]。多年生饲草玉米的亲本四倍体多年生大刍草茎节芽于1/2MS培养基培养时, 外源施加吲哚-3-丁酸(indole-3-butyric acid)并未影响茎节芽生根能力[18]。因此, 本试验各时期茎节扩繁系数的差异是否由上述激素引起值得研究, 并可能通过施加外源激素进一步提升多年生饲草玉米茎节扩繁系数。
多年生饲草玉米分株和茎节扩繁均可获得成功。茎节扩繁可提升建植成功率, 并不影响饲草产量。采用抽雄期中、下部长茎段育苗和覆膜越冬是释放多年生饲草玉米茎节扩繁潜力的有效措施。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|