{kind=link}
{kind=link}
{kind=link}
芒属植物分蘖数性状的QTL定位
[王延翠1, 2  , 于伟丽
, 于伟丽1, 2 , 王树楷1, 2 , 葛春霞1, 2 , 张国斌1, 2 , 陈翠霞1, 2, * ]
, 于伟丽]
|
|


作者简介:王延翠(1994-),女,山东聊城人,在读硕士。E-mail: wyc5515@163.com
芒属植物是一种多年生C4高大禾草,是一种重要的生物能源作物。分蘖是芒属植物重要的农艺性状之一,在调控其产量方面具有极其重要的作用。以五节芒和荻的种间杂交群体为材料,利用前期已构建的五节芒和荻的种间基因组遗传连锁图谱,结合2014年泰安、2015年泰安和东平3次重复调查的分蘖数表型数据,进行该重要性状的QTL遗传定位研究。结果表明:分蘖数频率分布呈现正态连续分布,符合数量性状遗传的特征;采用MQM复合区间作图法共定位到16个与分蘖数性状相关的QTL,单个QTL可解释的表型变异范围为11.4%~21.5%,LOD值为3.06~6.09。其中,3个QTL在3次定位分析中可重复检测到, qmfTI-2可分别解释12.7%、12.0%和15.5%的表型变异, qmsTI-1可分别解释12.0%、12.1%和19.8%的表型变异, qmsTI-2可分别解释21.5%、20.2%和13.4%的表型变异;3个QTL在2次定位分析中可重复检测到, qmfTI-1、 qmfTI-3和 qmsTI-4分别解释12.4%和11.4%、13.8%和13.2%、12.1%和14.3%的表型变异。通过对芒属植物分蘖数性状QTL分析,为芒属植物种质资源改良、分子标记辅助选择以及遗传学研究奠定基础。
Miscanthus is a tall, perennial plant of the grass family, with highly efficient C4 photosynthesis and therefore has potential to become an important bioenergy crop. Tiller number is one of important agronomic traits of Miscanthus and plays an extremely important role in yield regulation. We used two established interspecific genetic linkage maps of Miscanthus floridulus and M. sacchariflorus for quantitative trait locus (QTL) analysis of tiller number, together with the tiller number phenotype data from Tai’an in 2014, 2015 and Dongping in 2015. Results showed that the frequency of the tiller number phenotype followed the normal distribution of continuous variation, according with the hereditary characteristics of quantitative traits. A total of sixteen QTLs were identified for tiller number using a multiple-QTL model. The single QTLs explained 11.4%-21.5% of the phenotypic variation and the logarithm of odds (LOD) values of these QTLs ranged from 3.06 to 6.09. Three of the QTLs were detected repeatedly 3 times: qmfTI-2 explained 12.7%, 12% and 15.5% of the phenotypic variation, qmsTI-1 explained 12%, 12.1% and 19.8% of the phenotypic variation and qmsTI-2 explained 21.5%, 20.2% and 13.4% of the phenotypic variation. Three of the QTLs were detected twice, of which qmfTI-1, qmfTI-3 and qmsTI-4 explained, respectively, 12.4% and 11.4%, 13.8% and 13.2%, and 12.1% and 14.3% of the phenotypic variation. Through the QTL analysis of tiller number of Miscanthus, our study provides a good foundation for molecular marker-assisted breeding and improvement of Miscanthus germplasm resources.
芒属植物俗称芒草(Miscanthus spp.), 属于禾本科(Poaceae)、黍亚科(Panicoideae)、高粱族(Andropogoneae), 是一类C4多年生草本植物。芒属植物适应性强、生物质产量高、纤维质量好、燃值高, 作为可再生能源生产中的生物质源和造纸工业的原料得到了相当大的关注[1]。近年来, 许多国家加大对芒属植物的研究与开发[2], 一些欧美国家大规模推广种植[3]。芒属植物原产于亚洲的东部、东南亚、环太平洋群岛等地区, 中国是芒属植物的重要起源地之一, 有广泛的种质资源分布和遗传多样性[4], 目前世界上共有14个种, 在中国主要有7个种:芒(Miscanthus sinensis)、五节芒(M. floridulus)、荻(M. sacchariflorus)、南荻(M. lutarioriparius)、尼泊尔芒(M. nepalensis)、双药芒(M. nudipes)和红山茅(M. paniculatus)[1, 5]。相邻区域的芒属植物, 不同种类间及各种内、种间, 产生了大量的天然杂交种和丰富多样的不同地理生态类群[6], 这为芒属植物的遗传学和分子生物学研究提供了丰富的种质资源。
作为一种生物质能源植物, 芒属植物的育种目标之一是要提高产量, 从而获取更多的生物量。收获日期推迟到冬末、增加有效分蘖数等均可提高生物量[7]。分蘖是单子叶植物在生长发育过程中主茎基部形成的特化分枝, 并可以产生不定根。在水稻(Oryza sativa)、小麦(Triticum aestivum)等农作物中, 分蘖是主要农艺性状之一。分蘖数是影响水稻株型和产量的重要性状, Li等[8]克隆出水稻中第一个控制分蘖的基因MONOCULM1(MOC1), 而在对水稻产量相关基因的研究中发现, OsSPL14的过量表达植株表现为分蘖数减少、茎秆粗壮、抗倒伏能力增强, 植株产量增加[9]。在小麦中, 分蘖的多少会直接影响产量[10], 之前对小麦分蘖的遗传因子研究较多, 如tin、tin1、tin2及tin3等分蘖抑制基因[11, 12, 13, 14]。张倩辉等[15]研究发现, 小麦分蘖数受两对主效基因控制, 其中一对具有抑制分蘖的作用。多年生的芒属植物从初春开始分蘖, 以越冬茎为母茎, 由基部分蘖节处的侧芽开始第一次分蘖, 然后不断产生新的分蘖, 并在旗叶出现时达到高峰, 一直持续到深秋或初冬叶片枯萎时才停止分蘖[16]。由于芒属植物分蘖能力强, 分蘖数可达100以上[17], 分蘖相关的农艺性状对其产量有着主要的贡献。
分子生物学技术和基因组学的快速发展, 大大推动了作物的全基因组分子育种应用研究。然而, 迄今为止, 国内外关于芒属植物农艺性状QTL(quantitative trait locus, 数量性状基因座)鉴定的研究文献很少。作为一种新兴的草本能源植物, 芒属植物的遗传学及分子生物学研究远落后于玉米(Zea mays)、高粱(Sorghum bicolor)、水稻等作物。Atienza等[18, 19, 20, 21]利用随机扩增多态性DNA(random amplified polymorphic DNA, RAPD)标记, 构建了首张芒的遗传图谱, 并对产量、株高、花期和影响燃烧品质的矿物含量等性状进行了QTL定位分析。Liu等[22]以芒的两个栽培品种进行种内杂交, 构建遗传作图群体, 利用单核苷酸多态性(single nucleotide polymorphism, SNP)标记定位了3个斑马纹性状QTL和3个斑马纹强度QTL。Gifford等[23]利用846个单核苷酸多态性(single nucleotide polymorphism, SNP)和简单重复序列(simple sequence repeats, SRR)标记构建遗传图谱, 并对开花时间、株高、叶片宽度和产量等性状进行了QTL研究。Dong等[24]对连续调查两年的14个产量性状的表型数据进行分析, 并进行QTL定位。Ge等[25]利用五节芒和荻的高密度遗传连锁图谱对株高、抽穗时间和开花时间3个农艺性状进行QTL定位分析, 共鉴定出66个相关QTL。
芒属植物自交不亲和, 基因组高度杂合, 获得的F1代群体为分离群体, 可用作定位群体。本研究以中国芒属植物中二倍体五节芒和二倍体荻的种间杂交群体作为定位群体, 采用MQM(multiple-QTL model)复合区间作图法对芒属植物的分蘖数进行了QTL初步定位, 旨在了解芒属植物分蘖的遗传基础, 挖掘与分蘖数相关的QTL, 为挖掘候选基因、基因克隆和分子辅助育种奠定理论基础。
本试验以来自中国湖南的二倍体五节芒(A0430, MF)为母本, 以甘肃的二倍体荻(陕26, MS)为父本进行种间杂交, 创建了包含116个单株的F1定位群体。F1群体种子于2011年春季在温室播种。次年4月, 双亲和F1群体移栽到山东农业大学试验基地, 按株行距2 m× 2 m随机种植。2013年4月, 群体每个单株分取3个分蘖, 分别移栽到东平试验基地, 株行距为2 m× 2 m。
2014年对种植于山东泰安(117° 09' E, 36° 09' N)、2015年对种植于山东泰安和东平(116° 31' E, 35° 53' N)的双亲及群体的分蘖数进行调查统计, 每年11月上旬, 对每株F1个体的分蘖数进行计数。有3个完全展开叶的枝条为有效枝, 分蘖为个体有效枝的总数。
本研究采用双拟测交策略, 利用本实验室前期已构建好的五节芒和荻的遗传连锁图谱[25]为基础, 进行分蘖数的QTL定位。构建的五节芒的遗传图谱包含650个标记, 19个连锁群, 图谱总长为2053.31 cM, 标记间的平均遗传距离3.25 cM。荻的遗传图谱包含495个标记, 19个连锁群, 图谱总长为1684.86 cM, 标记间的平均遗传距离3.54 cM。用MapQTL6[26]软件, 采用MQM复合区间作图法, 定位该群体的分蘖相关QTL。进行作图分析时设置步长为1 cM, 在显著水平为0.02条件下, 进行1000次的排列检验。最后将LOD值≥ 3作为QTL存在的阈值。
Microsoft Excel 2010 软件用来对表型数据进行整理, 利用SPSS 17.0软件对亲本和F1群体表型进行差异显著性分析。变异系数计算公式:变异系数=(标准差/平均值)× 100%。
芒属植物是多年生植物, 需要一个较长的建立期。在欧洲国家田间试验一般被认为从第3年起已经完全建立[7, 27], 在预测育种的实际潜力时, 这一事实是至关重要的。本研究收集了2014年泰安、2015年泰安和东平的五节芒、荻及其F1群体的分蘖数据, 并进行统计分析(表1), 分析结果显示, 五节芒平均分蘖数为48.00, 荻平均分蘖数为169.67, 亲本五节芒和荻在分蘖数上具有极显著差异。F1群体的分蘖数在21~393, 3次调查数据的平均值分别为158.03、203.09和166.23。群体分蘖数性状的变异系数较大, 分别为35.67%、43.64%和30.46%, 说明五节芒和荻的种间杂交后代在该性状上具有较大的选择价值。根据F1群体中的偏度和峰度检验结果可以看出, 表型频率分布呈现正态连续分布, 同时, 分蘖数存在明显的超双亲分离现象, 说明分蘖数是由多个基因控制的数量性状, 适合进行QTL分析(图1)。
| 表1 亲本及其F1群体的分蘖数表型数据 Table 1 Phenotypic data for tiller number of parents and F1 population |
 | 图1 不同年份及地点的F1群体分蘖数的表型分布Fig.1 Frequency distribution of tiller number in the F1 population in different years and locations |
利用MQM复合区间作图法, 在五节芒图谱中定位到7个分蘖相关QTL, 分别在连锁群MF6、MF8、MF16和MF19, LOD值在3.06~6.00, 可解释的表型变异范围为11.4%~21.4%(表2和图2)。其中, qmfTI-2在3次定位分析中均能检测到, 位于第19连锁群, 可解释12.7%、12.0%和15.5%的表型变异; qmfTI-1和qmfTI-3重复检测到2次, 分别位于第8和6连锁群, 分别解释12.4%和11.4%、13.8%和13.2%的表型变异。在荻图谱中定位到9个分蘖相关QTL, 分别在连锁群MS7、MS8、MS16、MS18和MS19, LOD值在3.08~6.09, 可解释的表型变异范围为11.5%~21.5%(表2和图3)。其中qmsTI-1和qmsTI-2在3次定位分析中均能检测到, 分别位于第7和19条连锁群, 分别解释12.0%、12.1%、19.8%和21.5%、20.2%、13.4%的表型变异; qmsTL-5重复检测到2次, 位于第7连锁群, 可解释14.3%和12.1%的表型变异。其他QTL均仅能在单一环境下检测到。
| 表2 五节芒和荻中分蘖数性状相关QTL定位结果 Table 2 QTL analysis results of tiller number in M. floridulus and M. sacchariflorus |
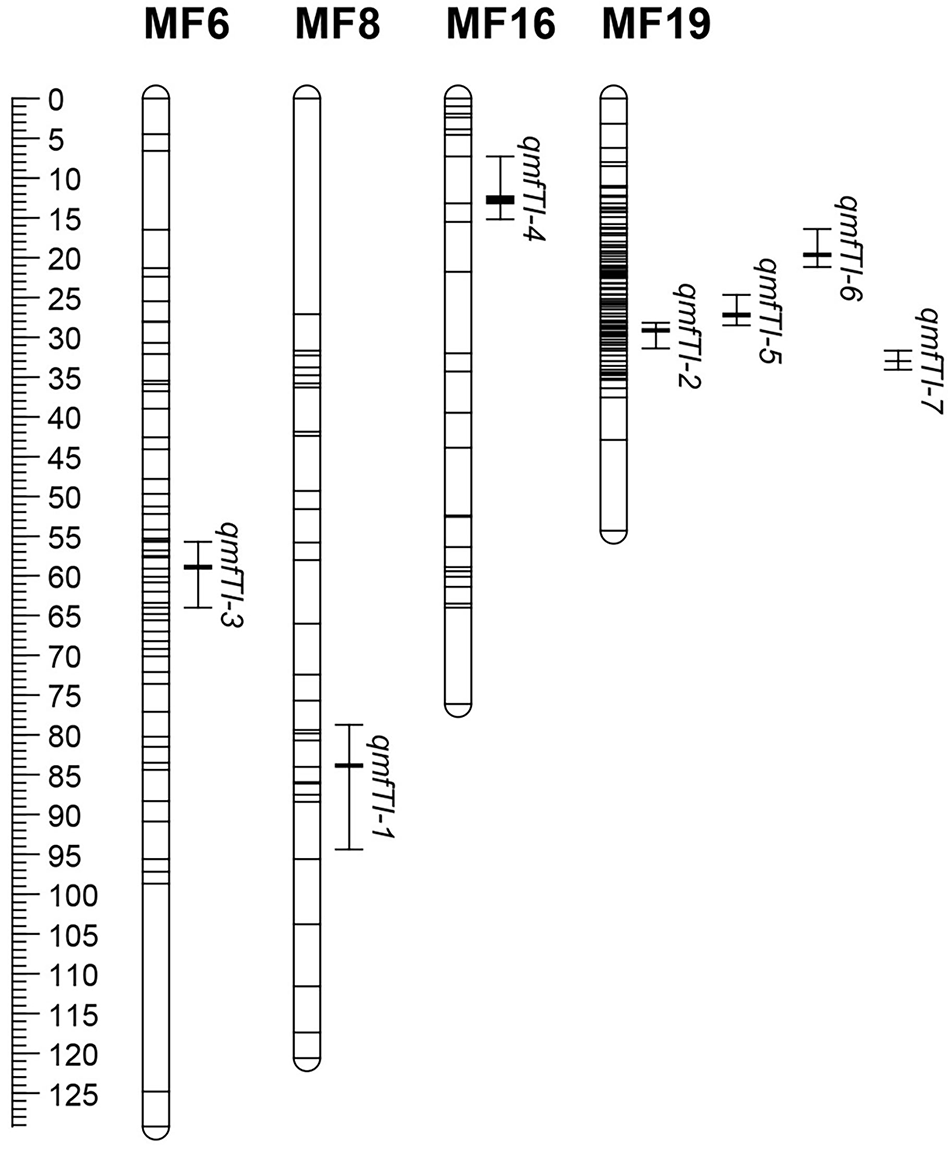 | 图2 分蘖数性状QTL在五节芒图谱上的分布Fig.2 Distribution of QTLs for tiller number on genetic map of M. floridulus |
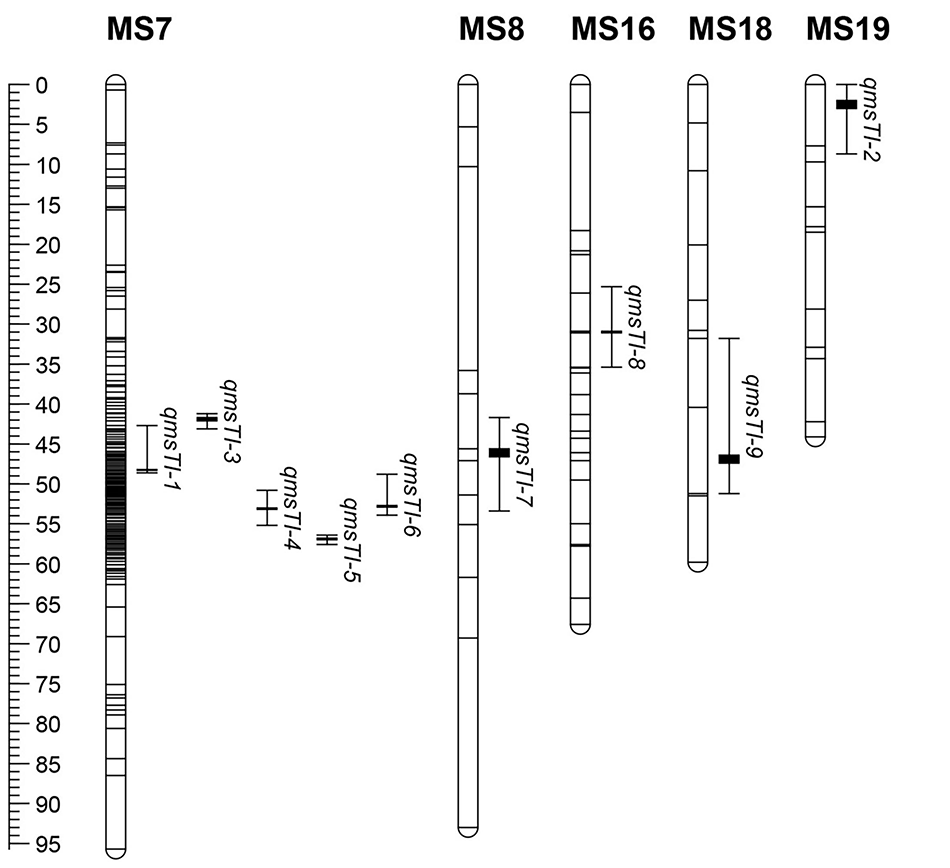 | 图3 分蘖数性状QTL在荻图谱上的分布 左边标尺显示以cM为单位的连锁群的长度。每个QTL中间的粗线为QTL的位置, 两边的线表示QTL的置信区间。下同。Fig.3 Distribution of QTLs for tiller number on genetic map of M. sacchariflorus The left ruler shows the length of the linkage group in centimorgan (cM). The thick line in the middle is the position of the QTL, and the lines on both sides represent the confidence interval of the QTL. The same below. |
在MF19连锁群24.702~34.129 cM检测到多个QTL, 同样在MS7连锁群41.205~57.564 cM也检测到了多个QTL的存在(图2和图3), Ge等[25]在对芒属植物株高、开花期和抽穗期等3个农艺性状进行QTL分析时, 发现在这两条连锁群上存在QTL簇, 物理位置分别是28.88~31.39 cM、46.62~52.54 cM, 本试验结果中的位置与其有重合, 进一步证实了这两个QTL簇的多效性。
分蘖性状显著影响分蘖作物的株型和生物产量。本研究双亲材料分别为来自湖南的五节芒和甘肃的荻, 五节芒在我国天然分布于长江中下游及以南区域, 具有植株高大、生物质产量高等特点[28, 29], 其根茎发达, 植株分蘖呈丛生状, 耐寒性差, 在山东不能越冬。荻分蘖呈散生, 分布范围广, 抗寒性强, 具有较强的生态适应性, 在山东地区生长良好[29, 30]。而五节芒和荻的杂交F1植株的分蘖性状, 介于双亲之间, 表现为紧凑型丛生和分散型丛生。在水稻、小麦、高粱和甘蔗(Saccharum officinarum)等作物中, 分蘖是影响产量的重要性状之一[9, 10, 31, 32]。在甘蔗中, 分蘖是其理想株型建成和合理群体结构形成的关键, 并显著影响宿根性, 进而影响产量[32]。对芒属植物来说, 分蘖性状包括分蘖数和分蘖角度等同样是影响产量的关键因素, Zhao等[33]在研究分蘖角度对产量的影响时认为, 紧凑型植株具有小的分蘖角度, 可在高密度种植模式下种植, 进而提高光合作用效率和生物产量。因此, 本研究针对分蘖性状, 利用全基因组作图的方法, 开展分蘖遗传定位研究。
芒属植物是一种高度杂合的异型杂交作物, 在F1代就会高度分离, 可利用F1代构建定位群体。艾辛等[34]用荻(B0134)和五节芒(A0430)的F1种间杂交群体进行分蘖数、株高等农艺性状的遗传分析, 研究结果显示, F1群体植株在分蘖性状中出现显著的超亲优势, 本研究与其结果一致。本试验结果显示, 两年两点共3次调查的F1群体表型均呈正态分布, 说明分蘖数是由多个基因控制的数量性状, 适合于QTL分析, 且所用的五节芒和荻遗传图谱总长分别为2053.31和1684.86 cM, 标记间的平均遗传距离分别为3.25和3.54 cM, 图谱质量高。
由于芒属植物目前没有全基因组信息, 国内外对芒属植物农艺性状QTL定位分析较少, QTL定位还处于初步阶段。对芒属植物进行QTL定位分析较多的性状主要是开花时间、株高和产量等[18, 19, 23, 25], 而对分蘖数的定位未有报道。Zhao等[33]对水稻和玉米中控制分蘖角度的基因TAC1的同源基因MsTAC1进行关联分析, 利用33个芒属植物种质鉴定出136个变异位点, 结果表明MsTAC1的变异影响分蘖角、分蘖数和生物量等性状。艾辛等[34]对荻与五节芒杂交F1代群体的主要农艺性状进行分析, 发现分蘖数与单株产量呈极显著相关, 说明分蘖数影响植株生物量。在另一种生物质能源植物柳枝稷(Panicum virgatum)的研究中, Das等[35]对柳枝稷产量相关农艺性状进行分析发现分蘖数性状与产量有着密切的正相关关系, 后期另一报道也证明了柳枝稷分蘖数以及分蘖生物量直接影响其生物产量[36]。本研究基于五节芒和荻为亲本创建种间杂交群体, 利用已构建好的两个亲本的高质量高密度遗传图谱, 首次对分蘖数性状进行QTL初步定位, 共定位到16个分蘖数相关QTL, 单个QTL最高可解释21.5%的表型变异, 其中3个QTL在3次定位分析中均能检测到, 3个QTL在2次定位分析中可检测到, 这6个QTL可以被认为是较稳定的QTL位点。
QTL簇影响多个农艺性状, 具有多效性, 在小麦、棉花(Gossypium spp.)、高粱等作物中普遍存在[37, 38, 39]。Ge等[25]在对芒属植物农艺性状相关QTL分析中, 发现了两个与株高、开花期和抽穗期相关的QTL簇, 分别在MF19(28.88~31.39 cM)和MS7(46.62~52.54 cM)连锁群上, 本研究中与分蘖数相关QTL有qmfTI-2和qmsTI-1、qmsTI-6分别存在于这两个QTL簇区域, 进一步证明了QTL簇的多效性, 也说明株高、抽穗期、开花期及分蘖数等性状之间存在相关性。本研究在MF19和MS7两个连锁群上分别定位到4和5个QTL, 而在其他连锁群上只定位到1个或没有定位到QTL, 虽然全基因组中标记总数多, 但在MF19和MS7两个连锁群上比较集中, 而其他连锁群标记较少, 这可能是本研究在这些连锁群上定位到的QTL数量较少或没有定位到的原因。其次, 作图群体的大小也影响QTL的定位, 有相关研究认为数量性状QTL定位分析群体至少要求500株[40]。有些QTL只检测到一或两次, 主要原因可能为:在同一生态点不同年限之间的气候如温度、降水和光照等差异较大, 个体的种植环境和表型鉴定时的人工误差等, 这些QTL稳定性较差, 其可靠性仍需进一步检验。
作为生物质能源植物, 产量是能源植物选择的重要标准, 提高产量是芒属植物育种的重要目标。其中分蘖相关性状对产量有着重要影响。本研究首次对分蘖性状进行QTL初步定位, 共定位到16个分蘖相关QTL, 这些QTL为芒属植物的MAS育种提供了重要的遗传信息。目前, MAS在玉米、水稻和小麦等作物育种中已得到广泛应用。分子标记技术与传统育种的有机结合, 使MAS广泛应用于玉米抗病、抗虫、抗旱、抗倒伏等育种工作中[41], 充分证明了MAS在实际生产应用中的重要性。张安宁等[42]通过MAS和常规回交育种相结合的方法, 将Bph6、Bph9、Bph14和Bph15等4个抗褐飞虱基因单独和聚合导入到节水抗旱稻恢复系中, 显著提高了节水抗旱稻恢复系的褐飞虱抗性。MAS同样在小麦的抗病和品质育种等方面得到有效利用[43, 44]。
芒属植物是多年生作物, 其繁殖方式一般为营养繁殖, 其实地试验的建立更是一项困难和昂贵的过程。例如, 在北欧国家, 芒属植物在移栽后的第一个冬季存活率较低, 实地试验的建立一般需要3年的时间[7], 这使其育种时间比一年生作物要长得多, 利用MAS可加快多年生作物的育种进程。本研究中检测到的QTLs是MAS育种的重要起点, 可以减少大量的育种时间。由于芒属植物高度自交不亲和, 传统的杂交育种和杂种优势育种方法存在较大的局限, 标记辅助选择育种是芒属植物育种的有效工具。本研究对芒属植物的分蘖性状进行QTL初步定位, 进一步了解了芒属植物分蘖的遗传基础, 对提高生物产量和种质资源改良具有重要意义。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|