
{kind=link}
{kind=link}
低温弱光胁迫下芸豆叶片光抑制与类囊体膜脂构成变化
引用本文
王玉萍, 郜春晓, 王盛祥, 何晓童. 低温弱光胁迫下芸豆叶片光抑制与类囊体膜脂构成变化. 草业学报, 2020,29(8): 116-125
WANG Yu-ping, GAO Chun-xiao, WANG Sheng-xiang, HE Xiao-tong. Changes in photoinhibition and fatty acid composition in the thylakoid membrane of kidney bean leaves under low temperature and weak light stress. Acta Prataculturae Sinica,2020,29(8): 116-125
Doi:10.11686/cyxb2020072WANG Yu-ping, GAO Chun-xiao, WANG Sheng-xiang, HE Xiao-tong. Changes in photoinhibition and fatty acid composition in the thylakoid membrane of kidney bean leaves under low temperature and weak light stress. Acta Prataculturae Sinica,2020,29(8): 116-125
Permissions
Copyright©2020, 草业学报编辑部
本文属于开放获取期刊。
低温弱光胁迫下芸豆叶片光抑制与类囊体膜脂构成变化
作者简介:王玉萍(1974-),女,甘肃天水人,博士,教授。 E-mail: wangyp@gsau.edu.cn
摘要
以对低温弱光敏感性有差异的两个红芸豆品种为材料,研究了低温弱光胁迫处理及恢复过程中幼苗叶片的叶绿素含量、荧光参数和类囊体膜脂肪酸组成的变化。结果表明:与对照相比,随胁迫时间延长,叶片中Chl a、Chl b和Chl(a+b)含量降低( P<0.05); Fv/ Fm、 Fv'/ Fm'、 qP、 ΦPSⅡ和ETR下降( P<0.05),Chl a/b和NPQ上升( P<0.05);类囊体膜脂MGDG、DGDG和SQDG中的亚麻酸(C18:3)含量显著降低( P<0.05),棕榈酸(C16:0)含量显著升高( P<0.05),PG中的棕榈酸(C16:0)和反式十六碳-烯酸[C16:1(3t)]含量降低( P<0.05),而亚麻酸(C18:3)和亚油酸(C18:2)含量升高( P<0.05),在叶片抵御低温弱光胁迫过程中维持一定的膜脂不饱和度的重要作用。随胁迫时间延长,类囊体膜总饱和脂肪酸(SFA)含量升高,多不饱和脂肪酸(PUFA)含量以及膜脂不饱和度(U/S)显著降低,恢复期则相反。在胁迫处理和恢复期,“英大红”和“小红芸豆”的U/S变化差异不明显,PUFA含量变化差异显著( P<0.05),且与 Fv/ Fm的相关性分别达到86.21%和83.92%,表明低温弱光处理及恢复过程中,光抑制后PSⅡ功能的修复与PUFA含量增加存在一定关系。因此,低温弱光胁迫下“英大红”光抑制程度较“小红芸豆”轻,可能是较高含量的PUFA增加了类囊体膜的不饱和度,维持膜的稳定性,减轻了光抑制。
关键词:
红芸豆; 低温弱光; 类囊体膜; 多不饱和脂肪酸; 叶绿素荧光
Changes in photoinhibition and fatty acid composition in the thylakoid membrane of kidney bean leaves under low temperature and weak light stress
Abstract
This research investigated the changes in photoinhibition and fatty acid composition in thylakoid membrane under combined stresses of low temperature and weak light in two varieties of kidney bean, ‘British big red’ and ‘Shanxi small red’, which have different sensitivity to low temperature and weak light. The main lipids in the thylakoid membrane are monogalactosyl diglyceride (MGDG), digalactosyl diglyceride (DGDG), phosphatidylcholine, phosphatidylglycerol (PG) and sulphoquinovosyl diglyceride (SQDG). The fatty acid composition of the thylakoid membrane, together with photosynthetic and chlorophyll (Chl) fluorescence parameters were determined during stress and recovery. It was found that with increase in stress duration, the content of Chl a, Chl b and Chl (a+b) decreased while the ratio of Chl a/b increased. Compared to the control, the maximum quantum yield of PSⅡ ( Fv/ Fm), the efficiency of excitation capture by open PSⅡ reaction centers ( Fv'/ Fm'), the photochemical quenching coefficient ( qP) and the actual PSⅡ efficiency ( ΦPSⅡ) declined, but Chl a/b and the non-photochemical quenching (NPQ) increased. With more prolonged stress duration MGDG, DGDG and SQDG, and thylakoid membrane content of linolenic acid (C18:3) were substantially reduced and the content of palmitic acid (C16:0) was concomitantly increased. These changes with increased stress duration represented an increased content of saturated fatty acids, and a decreased content of unsaturated fatty acids (UFA), polyunsaturated fatty acids (PUFA) and degree of membrane unsaturation in the thylakoid membrane lipids. However, there were also changes in the PG fraction which contributed to maintaining the overall degree of unsaturation in membrane lipids. This pattern was reversed during the recovery period. The correlation coefficients between PUFA and Fv/ Fm of ‘British big red’ and ‘Shanxi small red’ kidney beans were 0.8621 and 0.8392, respectively. Compared to ‘British big red’ kidney beans, stress caused more serious photoinhibition in PSⅡ of ‘Shanxi small red’ kidney bean leaves. Therefore, it can be postulated from these results that the reason for the increased the stability of the thylakoid membrane and reduced photoinhibition of PSⅡ in ‘British big red’ kidney beans may be the higher membrane content of UFA, especially PUFA.
Keyword:
red kidney bean; low temperature and weak light; thylakoid membrane; polyunsaturated fatty acids; chlorophyll fluorescence
温度和光照是影响农作物正常生长发育的重要因素。低温(零上低温)弱光是蔬菜生产中常常遇到的恶劣环境, 已经成为影响蔬菜产量和质量的重要因子。芸豆(Phaseolus vulgaris), 豆科菜豆属[1], 为我国重要的蔬菜作物。芸豆在我国多采用露地栽培, 尤其在冬春季和连绵阴雨天, 低温经常伴随着弱光发生, 对生长和产量造成严重影响。随着设施农业的发展, 芸豆的设施栽培面积逐年增加, 冬春季温室栽培常常伴有低温弱光逆境, 对生长和产量造成影响[2]。因此, 研究低温弱光对芸豆的生理特性影响, 对指导芸豆生产实践中正确选用优良品种和采取抗逆应变栽培技术具有重要的理论和实际意义。
类囊体膜由膜脂、膜蛋白及一些光合色素等成分组成, 低温能通过影响植物类囊体膜的结构而影响光合作用。在类囊体膜中, 主要存在单半乳糖甘油二酯(monogalatosyl diglyceride, MGDG)、双半乳糖甘油二酯(digalactosyl diglyceride, DGDG)、磷脂酰胆碱(phosphatidylcholine, PC)、磷脂酰甘油(phosphatidyl glycerols, PG)和硫代异鼠李糖甘油二酯(sulphoquinovosyl diglyceride, SQDG)等膜脂[3]。MGDG与PSⅡ 复合体外周天线相连, 而DGDG是PSⅡ 复合体的内部组分。SQDG与偶联因子CF0-CF1紧密结合, PG与捕光色素复合体II(light-harvesting complex II, LHCII)呈特异性结合[4]。植物对光能的吸收、传递和转换是在类囊体膜上结合的膜蛋白上进行的。低温胁迫会引起植物细胞结构发生变化, 细胞膜脂相变, 膜的流动性降低。膜脂组成影响膜的流动性, 从而影响膜通透性与膜蛋白的活性[5]。类囊体膜的流动性由膜脂组成及其不饱和度决定。研究发现, 类囊体膜脂不饱和度的增加能提高植物对低温的耐受性[6], 但是却增加植物对高温逆境的敏感性[7, 8]。低温胁迫下, 即使是中低等光强, 也会使植物发生光抑制甚至光氧化破坏[9]。因此, 光下低温比黑暗下低温更易使植物发生冷害, 低温与光照协同作用对植物的伤害更大。有关低温[6, 10], 高温[7, 8]以及盐胁迫下[11]植物极性脂和膜脂肪酸的变化已有较多报道, 然而有关低温与光照交互作用对类囊体膜脂不饱和度影响的研究较少[12], 尤其是低温弱光下芸豆类囊体膜脂肪酸组成与PSⅡ 功能关系的研究未见报道。
本试验在前期研究的基础上[13], 以对低温弱光敏感性存在明显差异的两个红芸豆品种为材料, 研究了低温弱光胁迫处理及恢复过程中两个品种幼苗叶片的叶绿素含量、荧光参数与类囊体膜脂肪酸的组成变化, 探索低温弱光胁迫下叶片光抑制与类囊体膜脂肪酸组成的关系, 为芸豆耐低温弱光育种材料的筛选以及冬春季温室高产栽培提供依据。
1 材料与方法
1.1 材料及处理
试验于2018年9月-2019年10月进行, 选择“ 英国大红芸豆” (简称“ 英大红” )和“ 山西小红芸豆” (简称“ 小红芸豆” )饱满种子在播种前晾晒1 d, 然后播种在装有营养土的10 cm× 10 cm的营养钵中育苗, 每钵2株。常规光温管理, 温度25 ℃/18 ℃(昼夜), 光周期12 h/12 h, 光强200 μ mol· m-2· s-1, 相对湿度75%。待幼苗长至34片真叶时, 选择长势一致, 生长健壮的植株移入人工气候箱进行处理。胁迫处理温度12 ℃/8 ℃(昼/夜), 光强80 μ mol· m-2· s-1, 光周期12 h/12 h。对照幼苗在温度为25 ℃/18 ℃(昼/夜), 光强200 μ mol· m-2· s-1, 光周期12 h/12 h下培养。分别在胁迫0、3、5、7 d及在正常温光下恢复1 (R1)、2 (R2)、3 d(R3) 时, 取第3片真叶进行叶绿素含量、荧光参数和类囊体膜脂肪酸含量测定。“ 英大红” 和“ 小红芸豆” 的处理组和对照组分别设为T和t, CK和ck。每处理10株, 3个平行。脂肪酸测定3次重复, 其余指标5次重复。
1.2 测定项目与方法
1.2.1 荧光参数测定 用FMS-2脉冲调制式荧光仪测定, 上午10:00-11:00进行。测定前先将叶片暗适应20 min, 测定初始荧光(minimum fluorescence, Fo), 再打饱和脉冲光测定最大荧光(maximal fluorescence, Fm), 打开光化光待荧光稳定后测得光适应下稳态荧光(steady-state value of fluorescence, Fs)、再打开饱和脉冲测得光适应下最大荧光(Fm') 以及最小荧光(Fo')等荧光参数, 光化学猝灭系数(photochemical quenching coefficient, qP)。根据公式计算:PSⅡ 最大光化学效率(maximum photochemical efficiency of PSⅡ , Fv/Fm), Fv/Fm=(Fm-Fo)/Fm、光化学淬灭系数, qP=(Fm'-Fs)/(Fm'-Fo'), PSⅡ 有效光能转化效率(effective photochemical efficiency of PSⅡ , Fv'/Fm'), Fv'/Fm'=(Fm'-Fo')/Fm', PSⅡ 实际光化学效率(actual photochemical efficiency of PSⅡ , Φ PSⅡ ), Φ PSⅡ =(Fm'-Fs)/Fm', PSⅡ 光合电子传递速率(electron transport rate, ETR), ETR=Φ PSⅡ × 0.42× PFD, 非光化学猝灭系数(non-photochemical quenching coefficient, NPQ), NPQ=(Fm-Fm')/Fm'[14, 15]。
1.2.2 叶绿素含量测定 采用丙酮浸提法测定[16]。洗净擦干叶片避开主脉剪碎混匀, 每个处理称取约0.2 g叶片放入试管中, 加入10 mL 80%丙酮避光提取48 h, 分别在 663和646 nm波长下测定吸光度。计算叶绿素a(Chl a)、叶绿素b (Chl b)和总叶绿素[Chl(a+b)]含量。
1.2.3 类囊体膜脂提取和分析 参照苏维埃等[17]的方法提取极性脂。按照朱亚芳等[18]的方法分析类脂。提取产物在薄层板(10 cm× 10 cm)上进行双向层析(thin layer chromatography, TLC), 所得类脂用苯∶ 石油醚=1∶ 1(V/V)提取, 再用400 mmol· L-1的氢氧化钾/甲醇甲脂化, 内标为花生酸甲脂(适量)。色谱条件:气相色谱, DB-23型30 m× 0.25 mm× 0.25 μ m石英毛细管柱; 柱温190 ℃, 进样口和检测器温度均为220 ℃; 载气为高纯氮, 氮气流速300 mL· min-1; 直接注射进样; 归一化法计算结果。
1.3 数据处理与分析
所有数据以平均值± 标准误(means± SD)表示, 用SPSS 19.0统计软件进行单因素方差分析, Duncan法进行差异显著性多重比较(P< 0.05)。用Origin 9作图。
2 结果与分析
2.1 低温弱光胁迫及恢复对叶绿素含量的影响
植物受到逆境胁迫时, 叶绿素代谢会发生变化。随着低温弱光胁迫处理时间延长, 两个品种红芸豆的叶绿素a (Chl a)、叶绿素b (Chl b)和总叶绿素Chl (a+b)含量均较对照降低(图1)。“ 英大红” 和“ 小红芸豆” 的Chl a含量在胁迫处理3、5、7 d分别较CK和ck降低了2.1%、4.2%、23.1%和8.0%、11.6%、27.1%, 恢复3 d分别较CK和ck低2.8%和6.4%。Chl b与Chl a的变化趋势相同, “ 英大红” 和“ 小红芸豆” 在胁迫3、5和7 d时Chl b分别较CK和ck降低4.3%、20.0%、39.0%和8.1%、22.6%、50.0%, 恢复3 d较CK与ck 低14.9%和25.8%, Chl b比Chl a恢复慢。Chl(a+b) 含量在胁迫处理过程中呈降低趋势, “ 小红芸豆” 的降低幅度较大, 恢复3 d仍与ck差异显著, 且胁迫处理和恢复过程中含量均低于“ 英大红” 。随胁迫时间延长, 两个品种的Chl a/b都呈上升趋势, 在胁迫57 d以及恢复3 d后的Chl a/b均高于相应的对照, 但是“ 英大红” 的升高幅度小而“ 小红芸豆” 的升高幅度大。因此, 低温弱光胁迫下“ 小红芸豆” 的叶绿素降解更严重, 恢复较慢。
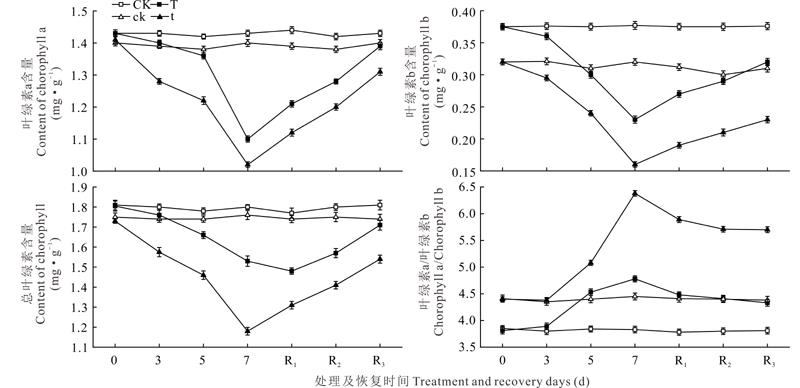 | 图1 低温弱光胁迫及恢复对红芸豆叶片叶绿素含量的影响 CK表示“ 英大红” 常规处理; ck表示“ 小红芸豆” 常规处理; T表示“ 英大红” 低温弱光及恢复处理; t表示“ 小红芸豆” 低温弱光及恢复处理。R1, R2, R3表示低温弱光处理后恢复的第1, 2, 3天。下同。Fig.1 Effect of stress and recovery under low temperature and weak light stress on chlorophyll content in the leaves of red kidney bean CK represent normal treatment for “ British red kidney bean ck represent normal treatment for “ small kidney bean” ; T represent low temperature and weak light treatment and recovery of "British red kidney bean” ; t represent low temperature and weak light treatment and recovery of “ small kidney bean” ; R1, R2, R3 represent the first, second and third recovery day after low temperature and weak light treatment. The same below. |
2.2 低温弱光胁迫及恢复对叶绿素荧光参数的影响
随胁迫处理延长, 两个品种的Fv/Fm、Fv'/Fm'、qP、Φ PSⅡ 和ETR均呈下降趋势, NPQ呈上升趋势, 恢复过程则相反(图2)。“ 英大红” 和“ 小红芸豆” 在处理3、5和7 d时Fv/Fm分别较CK和ck下降18.1%、26.0%、35.2%和22.0%、32.8%、49.8%, 恢复3 d分别较CK和ck低6.1%和15.2%。“ 英大红” 和“ 小红芸豆” 的Fv'/Fm'在胁迫处理3、5和7 d时分别较CK和ck降低了 6.9%、18.2%、28.0%和17.0%、26.6%、41.2%, 恢复3 d后分别较CK和ck低5.0% 和14.2%。“ 英大红” 和“ 小红芸豆” 的qP在胁迫处理3、5和7 d分别较CK和ck降低了16.8%、30.1%、46.2%和27.2%、46.1%、54.2%, 恢复3 d后较CK和ck低11.2%和19.8%。两个品种的Φ PSⅡ 在胁迫处理过程中呈下降趋势, “ 英大红” 和“ 小红芸豆” 在胁迫处理3、5和7 d分别比CK和ck下降了14.0%、36.4%、49.1%和18.1%、51.2%、69.1%, 恢复3 d后仍较CK和ck低12.0%和23.2%。“ 英大红” 和“ 小红芸豆” 的ETR在胁迫处理3、5和7 d较CK和ck降低了11.9%、33.1%、50.2%和19.1%、46.8%、70.0%, 恢复3 d后分别低于各自对照19.2%和24.0%。“ 小红芸豆” 和“ 英大红” 的NPQ在胁迫处理3、5和7 d较ck和CK升高20.2%、8.1%、35.0%和13.1%、43.2%、22.0%, 恢复3 d分别高于各自对照10.1%和6.0%。“ 小红芸豆” 的Fv/Fm、Fv'/Fm'、qP、Φ PSⅡ 及ETR与ck相比, 下降幅度较“ 英大红” 明显, 而NPQ上升幅度度大于“ 英大红” (图2), 恢复3 d仍与ck差异明显, 而“ 英大红” 的各指标在恢复3 d时基本接近CK。说明“ 英大红” 的PSⅡ 光抑制程度较“ 小红芸豆” 轻。
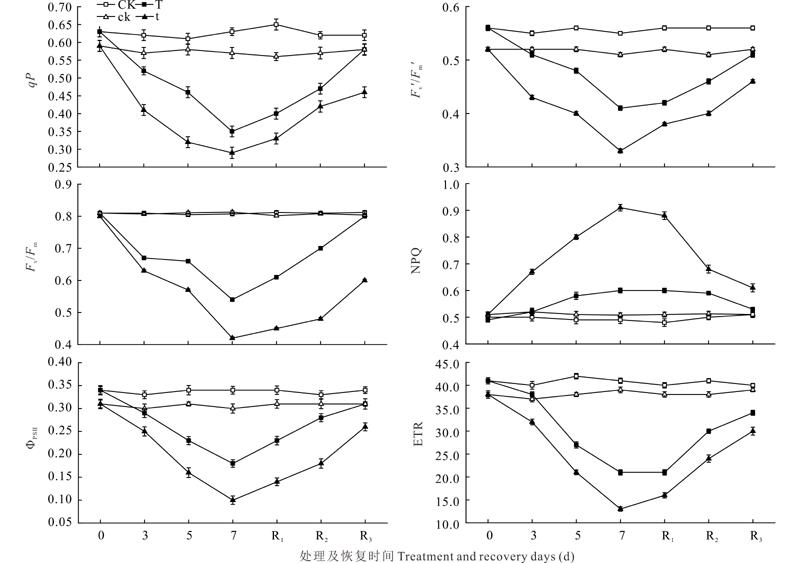 | 图2 低温弱光胁迫及恢复过程中红芸豆叶片叶绿素荧光参数的变化Fig.2 Changes of chlorophyll fluorescence parameters under low temperature and low light stress in the leaves of red kidney bean |
2.3 低温弱光胁迫及恢复过程中类囊体膜脂肪酸的组成变化
膜脂分析结果显示, 红芸豆类囊体膜脂包括MGDG、DGDG、SQDG和PG; 含有棕榈酸(C16:0)、反式十六碳-烯酸C16:1(3t)、硬脂酸(C18:0)、油酸(C18:1)、亚油酸(C18:2)和亚麻酸(C18:3), 每种脂肪酸在每种膜脂中的含量各不同(表1, 2)。随低温弱光处理时间延长, “ 英大红” 和“ 小红芸豆” 类囊体膜3种甘油酯MGDG、DGDG和SQDG中的亚麻酸含量与对照相比呈显著降低趋势(P< 0.05), 棕榈酸含量与对照相比显著升高(P< 0.05); MGDG中的亚油酸和硬脂酸含量显著升高(P< 0.05); SQDG中的亚油酸含量呈降低趋势(P< 0.05), 硬脂酸含量升高(P< 0.05); DGDG中油酸含量基本未变或稍有降低。恢复3 d后, “ 英大红” 和“ 小红芸豆” 类囊体膜上的DGDG和SQDG中的亚麻酸含量基本接近对照, 而MGDG中的亚麻酸分别低于对照4.39%和4.89%。“ 英大红” 类囊体膜3种甘油酯中亚麻酸的含量较“ 小红芸豆” 高, 恢复也快(表1, 2)。
| 表1 低温弱光胁迫处理及恢复对“ 英大红” 类囊体膜脂肪酸组成的影响 Table 1 Effect of fatty acid composition of thylakoid membrane lipids during low temperature and weak light treatment and recovery in “ British red kidney bean” (mol· mol-1) |
正常温光下, “ 英大红” 和“ 小红芸豆” 类囊体膜PG中总饱和脂肪酸[C16:0+C16:1(3t)+C18:0]含量分别为68.36%和69.83%(表1, 2)。随低温弱光胁迫时间延长, PG中总饱和脂肪酸相对含量降低(P< 0.05), 多不饱和脂肪酸(C18:3)和(C18:2)含量升高。胁迫处理35 d, “ 英大红” 类囊体膜PG中饱和脂肪酸含量下降(P< 0.05), 处理第7天有所回升, 但低于CK, “ 小红芸豆” 类囊体膜PG中总饱和脂肪酸含量与ck相比降低幅度小于“ 英大红” 。胁迫7 d, 两个品种PG中C16:0和C16:1(3t)相对含量下降, 恢复3 d与对照差异不显著, 而不饱和脂肪酸含量降至与对照基本相同。
| 表2 低温弱光胁迫处理及恢复对“ 小红芸豆” 类囊体膜脂肪酸组成的影响 Table 2 Effect of fatty acid composition of thylakoid membrane lipids during low temperature and weak light treatment and recovery in “ small kidney bean” (mol· mol-1) |
2.4 低温弱光胁迫及恢复对类囊体膜脂肪酸结构组成的影响
随低温弱光胁迫时间延长, 两个品种类囊体膜中的饱和脂肪酸(SFA)含量增加, 不饱和脂肪酸UFA(C18:1+C18:2+C18:3)、多不饱和脂肪酸PUFA(C18:2+C18:3)、不饱和指数(IUFA)以及膜脂不饱和度(U/S)均呈下降趋势, 恢复过程则相反; 在胁迫处理及恢复过程中, 单不饱和脂肪酸(MUFA)变化不大(表3)。“ 英大红” 类囊体膜PUFA和不饱和指数(IUFA)在胁迫处理及恢复过程中均高于“ 小红芸豆” , 但是U/S的变化差异不明显。相关分析表明, “ 英大红” 的PUFA与Fv/Fm的相关性达到86.21%(r=0.8621), “ 小红芸豆” 的PUFA与Fv/Fm的相关性为83.92%(r=0.8392), 表明光抑制后的修复与PUFA的恢复可能存在一定联系。
| 表3 低温弱光胁迫及恢复下红芸豆叶片类囊体膜脂肪酸结构组成分析 Table 3 Analysis of thylakoid membrane lipids structural group during low temperature and weak light treatment and recovery in red kidney bean (%) |
3 讨论
叶绿素在光合作用中有吸收、传递和转换光能的作用, 其含量的大小直接影响植物光合作用的强弱, 也是衡量植物耐逆境胁迫的重要生理指标之一。低温除了降低叶绿素总含量外, 也引起Chl a/b 值的变化[19]。Chl b主要位于PSⅡ 的捕光天线(LHCII), 而Chl a则主要存在于核心复合物中, 胁迫条件下Chl a/b变化能反映反应中心与LHCII对低温的敏感性和损伤程度[20]。本试验发现, 随低温弱光胁迫时间延长, 两个品种的Chl a, Chl b和Chl(a+b) 含量均较对照降低, 叶绿素发生部分降解; Chl a/b升高表明Chl b的降解程度大于Chl a, 说明LHCII对低温弱光的敏感性比反应中心蛋白高。与对照相比, “ 小红芸豆” Chl a/b升高幅度较“ 英大红” 大, 说明前者的LHCII对低温的敏感性较高。
低温胁迫下植物的CO2 同化受阻, 弱光下也会产生过剩的光能, 对光系统造成伤害[21]。Fv/Fm表示PSⅡ 最大光化学效率, Φ PSⅡ 代表PSⅡ 实际光化学效率, Fv'/Fm'为PSⅡ 激发能转化效率, qP为光化学猝灭系数, NPQ反映过剩光能的热耗散能力[22]。本研究表明, 低温弱光胁迫下红芸豆叶片Fv/Fm、Φ PSⅡ , Fv'/Fm'和qP较对照均显著降低(图2), PSⅡ 发生光抑制。“ 小红芸豆” 的Fv/Fm、Φ PSⅡ , Fv'/Fm'和qP的下降幅度较“ 英大红” 显著, 光抑制程度更严重。由于低温弱光下qP、Fv'/Fm'不断下降, PSⅡ 反应中心开放程度的降低以及天线捕光效率的下降都能导致用于光合电子传递的量子产量减少, 使Φ PSⅡ 降低。ETR反映PSⅡ 向PSI的线性电子传递, 与碳同化能力大小有关[22]。本试验表明, 随低温弱光胁迫时间延长ETR降低显著, 可能与低温弱光抑制碳同化有关, 即逆境下由于qP减小, 受体QA重新氧化能力减弱, PSⅡ 电子传递活性降低; 与对照相比, 虽然“ 小红芸豆” 的ETR较“ 英大红” 降低显著, 但是NPQ升高幅度大于“ 英大红” , 说明LHCII吸收的光能热耗散增加, 用于光化学反应的光能减少, CO2同化力下降, PSⅡ 通过光抑制响应低温弱光进行保护。
脂肪酸是细胞膜脂的构成组分, 低温胁迫诱发活性氧造成不饱和脂肪酸双键断裂, 发生膜脂过氧化, 使不饱和脂肪酸含量降低[20, 23], 且脂类分子的不饱和程度越高, 氧化作用发生越明显[24]。本研究发现, 低温弱光胁迫下红芸豆类囊体膜3种甘油酯中的亚麻酸含量降低幅度最大, 棕榈酸含量升高最明显, 其余脂肪酸升降程度不同, 因此类囊体膜脂总饱和度增加, 是不同膜脂中的脂肪酸变化的综合结果, 与代玉华等[25]在黄瓜(Cucumis sativus)上的研究结果相同。本试验中, 单不饱和脂肪酸含量在胁迫处理与恢复过程中变化不明显, 表明其稳定性大于多不饱和脂肪酸。尽管膜饱和度增加有利于降低膜脂过氧化, 然而一定程度的膜脂不饱和度是维持膜流动性及其功能的基础[24]。PG作为影响类囊体膜脂发生相变的关键因素, 不饱和脂肪酸含量增加有利于低温下膜稳定性与流动性的保持[26, 27]。本研究发现, 随低温弱光胁迫时间延长, PG中棕榈酸和反式十六碳-烯酸的含量降低(P< 0.05), 亚麻酸和亚油酸含量升高(P< 0.05), 在红芸豆抵御低温弱光胁迫中起维持一定的膜脂不饱和度的重要作用。PG中饱和脂肪酸反式十六碳-烯酸的含量与LHCII的结构有一定关系[20], 含量降低使LHCII寡聚体解聚, 捕光效率下降[28]。低温弱光胁迫下, “ 英大红” PG中的反式十六碳-烯酸的含量降低程度较“ 小红芸豆” 小, LHCII结构较稳定, 捕光效率较高。但是恢复过程中两个品种PG中饱和脂肪酸含量一直处于升高趋势, 与温度降低致膜脂相变相反。然而, Fv/Fm较快恢复说明, PG中饱和脂肪酸含量的增加并没有影响到PSⅡ 光抑制的修复。有研究表明, 光抑制的恢复需要PG中含有一定量的饱和脂肪酸利于降低膜脂过氧化, 但是膜脂过饱和化则会导致膜功能受损, 限制光抑制的修复[29]。因此, 恢复过程中红芸豆类囊体膜PG中饱和脂肪酸含量的增加可能在限定范围内, 对光抑制后恢复的影响作用不大。
随低温弱光胁迫时间延长, 红芸豆类囊体膜脂肪酸的组成变化结果趋向于饱和化, 但是PG中亚麻酸和亚油酸含量升高, 维持膜脂不饱和度的作用, 对于降低膜脂相变和低温弱光敏感性起重要作用。“ 英大红” 类囊体膜中不饱和度(U/S)和不饱和指数(IUFA)在胁迫处理及恢复过程中均高于“ 小红芸豆” 。在胁迫及恢复过程中, 尽管两个品种U/S变化差异不大, 但是多不饱和脂肪酸(PUFA)含量变化差异显著(P< 0.05)。说明在恢复过程中PSⅡ 光抑制的修复与PUFA的关系较与膜脂U/S的关系密切。有研究表明, 低温下烟草(Nicotiana tabacum)光抑制加重并且恢复减慢的重要因素是PUFA含量下降所致[30]。进一步通过相关性分析发现, 低温弱光下红芸豆类囊体膜中PUFA与Fv/Fm变化均具有较高相关性, 表明光抑制的修复与PUFA含量升高存在一定联系。PSⅡ 光抑制的特征是反应中心D1蛋白周转加快[31], 类囊体膜脂不饱和度降低有可能减慢了重新合成的D1蛋白装配到PSⅡ 反应中心的速率[32]。“ 英大红” 类囊体较高含量的PUFA增加了膜脂不饱和度, 维持了类囊体膜功能, 减轻了PSⅡ 光抑制。
4 结论
低温弱光胁迫处理与恢复过程中, PSⅡ 光抑制后的修复与PUFA含量增加存在一定关系。低温弱光胁迫下“ 英大红” 光抑制程度较“ 小红芸豆” 轻, 可能是由于较高含量的PUFA增加了类囊体膜不饱和度, 维持膜生理功能, 降低了PSⅡ 对低温弱光的敏感性。因此, 生产实践中可以通过物理或者化学方法增加芸豆类囊体膜脂中不饱和脂肪酸含量, 降低PSⅡ 的光抑制和光氧化, 提高作物对低温弱光的耐受性。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|