




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
酸胁迫对夏枯草叶绿素荧光特性和根系抗氧化酶活性的影响
[张利霞1 , 常青山2, *  , 薛娴
, 薛娴1 , 刘伟1 , 张巧明2 , 陈苏丹2 , 郑轶琦2 , 李景林2 , 陈婉东2 , 李大钊3 ]
, 薛娴|
|


作者简介:张利霞(1982-),女,河南荥阳人,讲师,博士。E-mail: hkdzlx@126.com
为了阐明夏枯草幼苗对酸性环境的耐受性,采用不同pH值(pH=4.5、4.0、3.5、3.0)的Hoagland营养液模拟酸胁迫环境,研究酸胁迫对夏枯草幼苗叶绿素荧光特性及根系抗氧化酶活性的影响。结果表明,随着pH值下降,代表供体与受体侧参数的K点相对可变荧光( WK)、J点相对可变荧光( VJ)和电子受体
, XUE XianThis research aimed to elucidate the tolerance of Prunella vulgaris seedlings to acid environments. Hoagland nutrient solutions with a range of pH values (pH=4.5, 4.0, 3.5, 3.0) were used to simulate environmental acid stress, and the effects of acid stress on chlorophyll fluorescence characteristics and root antioxidant activity were studied. It was found that at lower pH the quantum efficiency of PSⅡ electron transfer ( φEo) in leaves of P. vulgaris decreased, while a number of other fluorescence parameters increased, including: the variable fluorescence Fk to the amplitude Fj- Fo ( WK), relative variable fluorescence at J step ( VJ),
近年来, 随着世界经济发展、工业化进程加快, 酸雨污染问题日益严重, 其对生态系统及人类生活环境已造成严重影响, 成为全球范围内亟待解决的重要环境问题之一[1]。中国近年来酸雨发生面积不断扩大, 几乎成燎原之势, 继欧洲和北美之后, 跻身世界第三大酸雨区[2]。我国强酸雨区的面积较大, 长江以南地区是我国主要的酸雨控制区[3]。植物是陆地生态系统中酸雨污染最直接的受体, 酸雨对植物生长发育的作用机理及植物对酸雨的抗性机制一直以来都是研究的热点[4]。研究表明, 酸雨直接通过积累活性氧、破坏细胞质膜与叶绿体超微结构、使植物酶失活、降解叶绿素、损坏光合系统, 从而导致植物生长发育不良或畸形[5]。同时, 酸雨沉降至地面, 会导致土壤酸化, 抑制土壤呼吸和植物根系的生长, 加重土壤养分流失, 从而使植物根毛对水分和养分的吸收能力显著下降[6], 造成植物的生长发育受到抑制, 并导致农作物的产量与品质下降。尤其是在幼苗期, 酸雨对植物生长发育的抑制作用尤为显著[7]。
夏枯草(Prunella vulgaris)是唇形科夏枯草属的多年生草本植物, 因其果穗具有重要的功效与药理作用, 受到了人们青睐, 在临床上被广泛用于治疗高血压、糖尿病、癌症、肺结核和肝炎等重大疾病[8, 9, 10]。夏枯草资源主要分布于长江中下游地区, 主产区集中分布于江苏、安徽、河南、江西和浙江等地区, 这些地区多是酸雨发生较多的地区[2, 11]。近年来, 夏枯草野生资源急剧减少, 其人工栽培种植是保障夏枯草资源可持续发展的重要举措。目前, 关于环境因子对夏枯草生理特性、产量及品质影响的研究主要集中于水分、养分、光照等因子的调控作用[12, 13], 而对于酸胁迫如何影响夏枯草的内部光系统以及根系抗氧化酶的机制尚不清楚。因此, 本研究通过设置不同pH值营养液, 模拟土壤酸性环境, 研究苗期夏枯草叶绿素荧光特性及根系保护酶活性对酸胁迫环境的响应特征, 以期为阐明夏枯草对酸性环境的耐受机制提供基础数据。
2018年10月进行夏枯草育苗:将夏枯草种子(采于河南驻马店)消毒后, 播种于装有等量营养土基质的周转箱(60 cm× 42 cm× 15 cm)中进行育苗, 正常肥水管理。次年, 挑选长至4叶期且长势均衡的幼苗, 于自然光照, 温度为昼夜(30± 3) ℃/(20± 3) ℃的条件下, 放入装有1/2 Hoagland营养液的周转箱中进行泡沫板悬浮培养, 每隔34 d换一次营养液。
待夏枯草幼苗长到68叶期时, 选择大小相当的幼苗进行酸胁迫试验, 每个处理30株。根据前期预实验结果对夏枯草幼苗进行相应的酸胁迫处理。用硫酸和硝酸的混合液(按摩尔浓度5∶ 1)调制到相应pH值配制模拟酸胁迫母液, 与1/2Hoagland营养液混合配制成pH分别为4.5, 4.0, 3.5, 3.0的模拟酸溶液, pH 6.5为不含酸液的1/2Hoagland营养液。每个处理的营养液均每3 d更换一次。连续胁迫处理7 d后进行夏枯草荧光与生理指标的测定。
快速荧光诱导动力学参数的测定:夏枯草叶片经过充分暗适应后, 利用多功能植物效率分析仪(M-PEA, Hansatech, 诺福克, 英国)经3000 μ mol· m-2· s-1的脉冲光诱导, 测定夏枯草叶片快速叶绿素荧光诱导动力学曲线, 根据文献[14]方法计算JIP-test参数:光系统Ⅱ (photosystem Ⅱ , PSⅡ )最大光化学效率(Fv/Fm)、以吸收光能为基础的光合性能指数(performance index on absorption basis, PIabs)。PSⅡ 电子传递的量子效率(quantum efficiency of PSⅡ electron transfer, φ Eo)、可变荧光Fk占Fj-Fo振幅的比例(ratio of the variable fluorescence Fk to the amplitude Fj-Fo, WK)、J点的相对可变荧光(relative variable fluorescence at J step, VJ)、电子受体
采用氮蓝四唑(nitro-blue tetrazolium, NBT)还原法测定幼苗根系超氧化物歧化酶(superoxide dismutase, SOD)活性, 采用愈创木酚法测定根系过氧化物酶(peroxidase, POD)活性, 利用分光光度计法测定根系抗坏血酸过氧化物酶(ascorbic acid peroxidase, APX)活性, 采用蒽酮硫酸法测定叶片可溶性糖含量, 采用磺基水杨酸法测定叶片脯氨酸含量[15]。每个处理重复3次, 取平均值。
采用Excel 2007录入数据并计算平均值和标准差, 利用SPSS 16.0进行多重比较(Duncan)分析, P< 0.05表示差异显著。
随着pH值的降低, 反应中心吸收的光能用于电子传递的量子效率(φ Eo)呈逐渐下降的趋势, 在pH 4.5和4.0时低于对照, 但差异不显著(图1)。在pH 3.5和3.0时显著低于对照(P< 0.05), 说明酸胁迫处理不同程度地降低了光系统Ⅱ (PSⅡ )的相对电子传递能力。在pH 4.53.5时, 可变荧光Fk占Fj-Fo振幅的比例(Wk)高于对照, 但无显著差异; 只有在pH为3.0时, Wk显著高于其他处理(P< 0.05), 这表明放氧复合体(oxygen evolving complex, OEC)可能在pH 3.0处理下受到显著伤害。
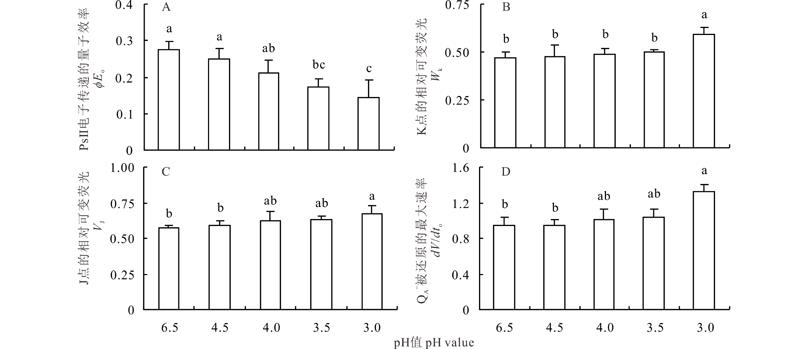 | 图1 酸胁迫下夏枯草叶片PSⅡ 电子传递的量子效率、K点的相对可变荧光、J点的相对可变荧光和QA-被还原的最大速率的变化 不同小写字母表示各处理在P< 0.05水平上差异显著, 下同。Fig.1 Changes of chlorophyll fluorescence parameters of quantum efficiency of PSⅡ electron transfer (φ Eo), ratio of the variable fluorescence Fk to the amplitude Fj-Fo (Wk), relative variable fluorescence at J step (VJ) and QA- maximum rate of reduction (dV/dto) in P. vulgaris leaves under acid stress Different lowercase letters indicate significant differences in the level of P< 0.05 for each treatment, the same below. |
VJ反映了J点的相对可变荧光强度, 可以表征QA-的积累量。随着pH值的降低, VJ值呈逐渐增加趋势, 在pH 3.0处理下显著高于对照(P< 0.05)。dV/dto反映QA-被还原的最大速率, dV/dto在pH 4.53.5处理下与对照差异不显著, 在pH 3.0处理下显著高于对照(P< 0.05)。pH 4.53.5处理下VJ与dV/dto的上升幅度较小, 表明电子传递受到影响较小, 而pH 3.0处理下两者上升幅度较大, 说明该处理下电子传递能力受到显著抑制。
酸胁迫增加了夏枯草叶片单位反应中心吸收能量(ABS/RC), pH 4.03.0处理与对照差异显著(P< 0.05), pH 3.0处理上升幅度最大(图2)。单位反应中心捕获能量(TRo/RC)在pH 4.53.5处理下逐渐增加, 但与对照差异不显著, 在pH 3.0处理下上升幅度较大, 且显著高于对照(P< 0.05)。夏枯草叶片单位反应中心传递的能量(ETo/RC)总体呈降低趋势, 各处理与对照相比均未达显著差异。随着pH值的降低, 夏枯草叶片单位反应中心的热耗散(DIo/RC)呈逐渐升高的趋势, 其中pH 4.5处理与对照差异不显著, 其他处理均显著高于对照(P< 0.05)。
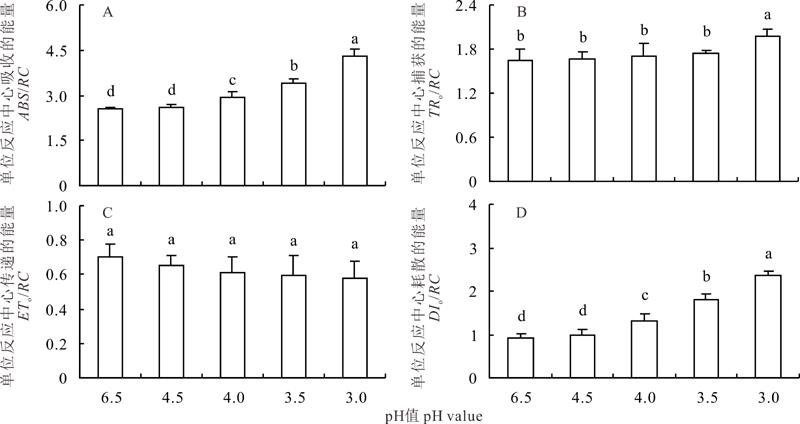 | 图2 酸胁迫下夏枯草叶片单位反应中心吸收、捕获、用于电子传递和热耗散的能量变化Fig.2 Changes of absorption flux per reaction center (ABS/RC), trapped energy flux per RC (TRo/RC), electron transport flux per RC (ETo/RC) and dissipated energy flux per RC (DIo/RC) in P. vulgaris leaves under acid stress |
Fv/Fm, 是指光合机构把吸收的光能用于光化学反应的最大效率。随着pH值的降低, 夏枯草Fv/Fm呈下降的趋势, pH 4.5处理与对照差异不显著, 而pH 4.03.0处理均显著低于对照(图3)。基于光吸收性能的指数(PIabs)也呈逐渐降低趋势, 除pH 4.5处理与对照差异不显著外, 其他处理均显著低于对照(P< 0.05)。
 | 图3 酸胁迫下夏枯草叶片最大光化学效率和光合性能指数的变化Fig.3 Changes of the maximum quantum yield of PSⅡ (Fv/Fm) and performance index on absorption basis (PIabs) in P. vulgaris leaves under acid stress |
随着pH值的降低, 夏枯草叶片光系统Ⅰ (PSⅠ )的最大氧化还原活性(Δ I/Io)呈降低趋势, pH 4.53.5处理与对照差异不显著, 而pH 3.0处理显著低于对照(P< 0.05), 但与pH 3.5差异不显著(图4)。酸胁迫降低了夏枯草叶片PSⅡ 与PSⅠ 之间的协调性(Φ PSⅠ /PSⅡ ), 其中pH 4.53.5处理与对照差异不显著, 而pH 3.0处理显著低于对照及其他处理(P< 0.05)。说明酸胁迫不但降低了夏枯草叶的PSⅠ 的活性, 也破坏两个光系统之间的协调性(Φ PSⅠ /PSⅡ )。
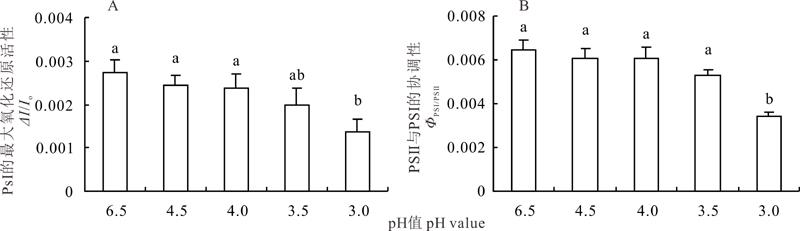 | 图4 酸胁迫下夏枯草叶片PSⅠ 的最大氧化还原活性及叶片PSⅡ 与PSⅠ 的协调性的变化Fig.4 Changes of the maximum PSⅠ redox acitity (Δ I/Io) and the coordination of photosystem between Ⅰ and Ⅱ (Φ PSⅠ /PSⅡ ) in leaves of P. vulgaris under acid stress |
随着pH值的降低, 夏枯草幼苗根系超氧化物歧化酶(SOD)活性呈逐渐下降的趋势(表1)。与对照相比, 不同酸胁迫处理下根系SOD活性均显著降低, 且不同酸处理之间差异显著(P< 0.05)。根系过氧化物酶(POD)活性的变化趋势和SOD活性相类似, 随着pH值下降逐渐降低, 各酸胁迫处理均显著低于对照(P< 0.05)。随着pH值的降低, 根系抗坏血酸过氧化物酶(APX)活性呈先上升后下降的趋势, 但均显著高于对照, 在pH值为3.5时达到最大值, 显著高于其他处理(P< 0.05)。
| 表1 不同pH值对夏枯草幼苗根系超氧化物歧化酶、过氧化物酶和抗坏血酸过氧化物酶活性的影响 Table 1 Effects of different pH values in nutrient solutions on activities of superoxide dismutase, peroxidase and ascorbic acid peroxidase in roots of P. vulgaris seedlings (U· g-1· min-1) |
夏枯草叶片中可溶性糖含量随着营养液pH值的降低, 呈逐渐上升的趋势, pH值为4.53.5时, 可溶性糖的含量高于对照, 但差异不显著(图5), 在pH值为3.0时达到最大值, 且显著高于对照(P< 0.05)。叶片中脯氨酸的含量变化趋势与可溶性糖含量势相似, 随着营养液pH值的降低呈逐渐上升趋势, pH为4.5时, 与对照无显著差异, pH为4.03.0时, 均显著高于对照。
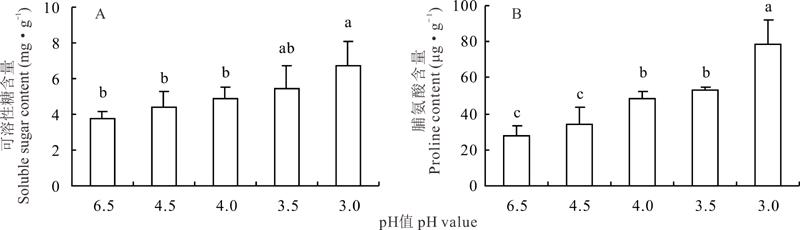 | 图5 不同pH值对酸胁迫下夏枯草可溶性糖与脯氨酸含量的影响Fig.5 Effects of different pH values in nutrient solutions on the contents of soluble sugar and proline in leaves of P. vulgaris under acid stress |
叶绿素荧光诱导动力学参数能够用于描述植物光合作用的机理及光合生理状况, 可作为检测植物遭受胁迫伤害程度的指标[16]。有研究表明, PSⅡ 在逆境条件下因受胁迫活性下降, 线性电子传递链在传递电子受阻后会导致活性氧的产生与积累[17]。本研究中, φ Eo不断下降, 说明PSⅡ 的相对电子传递能力下降; VJ与dV/dto均逐渐升高, 表明QA到QB之间的电子传递受阻[18], 在pH 3.0酸胁迫下的φ Eo显著低于对照, 而VJ和dV/dto显著高于对照, 表明该胁迫下PSⅡ 的电子传递能力严重受阻, 在QA到QB电子传递受到的抑制较重, 并导致QA-的明显积累。Wk, 即K点相对可变荧光, 被人们广泛用于分析逆境胁迫对光合机构PSⅡ 供体侧, 即放氧复合体(OEC)伤害的程度[19]。在本试验中, Wk随着酸胁迫程度加重而逐渐增加, 表明夏枯草叶片PSⅡ 放氧复合体(OEC)受到了伤害。pH 3.0处理下的Wk的上升幅度高于VJ, 表明该处理下PSⅡ 供体侧伤害程度重于受体侧[20]。酸胁迫下夏枯草叶片的PSⅡ 性能降低, 必然导致其向PSⅠ 传递电子的能力下降。
ABS/RC用来衡量PSⅡ 反应中心捕光天线色素的大小[21], 该指标升高说明叶片捕光天线色素增加, 意味着部分PSⅡ 反应中心的失活。ABC/RC和TR/RC增加, 说明叶片通过增加捕光天线色素的大小以及对光的捕获能力, 从而增强单一反应中心对光的吸收与捕获能力。夏枯草叶片单位面积的热耗散(DIo/RC)在酸胁迫下升高, 用于电子传递的能量(ETo/RC)减少, 说明叶片减少用于电子传递的量子份额, 通过增加热耗散来将过多的光能耗散掉, 来减少光抑制对叶片产生的伤害。Fv/Fm和PIabs两个指标反映了植物叶片PSⅡ 反应中心活性。在本研究中, φ Eo下降, VJ、dV/dto、Wk与ABS/RC升高, 说明酸胁迫导致PSⅡ 反应中心失活, PSⅡ 电子传递受阻, 进一步造成了Fv/Fm的降低, 夏枯草叶片发生了光抑制, 导致其叶片光能利用率降低[22]。PIabs, 光化学性能指数, 能从光能吸收、捕获和电子传递3个方面内容来综合反映PSⅡ 的活性[23]。本试验中, 由于PSⅡ 的结构和功能受到破坏, 因此导致夏枯草叶片PIabs的下降, 且叶片PIabs的下降幅度明显大于Fv/Fm的下降幅度, 说明PIabs对酸胁迫的反应比Fv/Fm更为灵敏, 本研究与前人研究一致[16]。
Δ I/Io表征的是PSⅠ 反应中心的最大氧化还原能力, 可以用来评价植物叶片PSⅠ 性能, 而Φ PSⅠ /PSⅡ 表征的是两个光系统之间的协调性[24]。本试验研究表明, pH 3.0处理下夏枯草的Δ I/Io显著下降, 说明该胁迫条件下PSⅠ 性能受到显著伤害, 这将抑制电子流从PSⅡ 流向PSⅠ , 引起QA-大量积累, 阻碍QA-之后的电子传递。因此, PSⅡ 与PSⅠ 之间电子传递受阻, 导致pH 3.0处理下的PSⅡ 与PSⅠ 之间的协同性(Φ PSⅠ /PSⅡ )明显失衡, 这可能是引起QA后续电子传递链效率下降以及整个电子传递链功能降低的主要原因[25]。
酸胁迫会诱导作物体内产生大量活性氧(ROS), 过量活性氧会破坏细胞膜的结构与功能, 导致细胞生理功能紊乱[26, 27]。植物在遭受酸胁迫时, 会通过维持几种抗氧化酶之间的协调和平衡来提高植株抗活性氧的能力[28]。本研究中酸胁迫下夏枯草根系的SOD与POD活性不断下降, 可能是由于根系在酸胁迫下产生了大量的活性氧并超出了根系SOD与POD的清除能力, 致使根系SOD、POD活性下降; 这与平邑甜茶在酸化处理一定天数后根系SOD与POD活性下降[27]的结果相似, 类似的结果也出现在萝卜(Raphanus sativus)与莴笋(Lactuca sativa)的相关研究中[29]。APX参与抗坏血酸清除过氧化氢的过程[16], 夏枯草根系APX活性在酸胁迫处理下均表现出高于对照的活性, 这说明夏枯草在酸胁迫条件下可以通过增强根系APX活性, 增强植株根系清除过氧化氢的能力, 缓解ROS对植物根系的伤害, 提高植株抵抗酸胁迫的能力。
前人研究表明, 当OEC活性受到抑制时会造成放氧过程中水的不完全裂解, 导致H2O2的产生; PSⅡ 受体侧电子传递受阻会使过剩电子泄露, 细胞中游离的O2因受泄露的电子攻击而生成超氧阴离子, 这些活性氧与超氧阴离子会造成细胞膜质过氧化增加和电解质渗漏等问题[30, 31]。本研究表明, 酸胁迫处理导致夏枯草叶片的OEC受到伤害, 电子传递受阻, 因此产生的活性氧造成夏枯草叶片细胞膜受到不同程度的伤害。在逆境胁迫条件下, 植物能够通过增加可溶性糖、脯氨酸等渗透调节物质的含量来适应逆境过程[32, 33]。本试验发现夏枯草叶片通过提高酸胁迫条件下的脯氨酸与可溶性糖含量, 来调节细胞液渗透压, 保护细胞膜的稳定性。特别地, 脯氨酸对羟基自由基清除和保护酶也具有一定的保护和稳定作用[34], 脯氨酸增幅大于可溶性糖增幅, 说明脯氨酸可能已成为主要的渗透调节物质[35]。由此说明, 夏枯草在酸胁迫条件下, 通过增加可溶性糖、脯氨酸的积累来提高细胞液的浓度, 维持细胞的水势, 与根系的抗氧化酶协同作用, 清除活性氧, 保护光系统和增加细胞膜的稳定性, 以减少酸雨带来的伤害。
综上所述, 酸胁迫造成放氧复合体(OEC)受到伤害, PSⅡ 的光合电子传递受阻, 抑制了PSⅡ 与PSⅠ 的活性, 降低了PSⅡ 与PSⅠ 的协调性, 导致夏枯草SOD与POD抗氧化酶活性下降。夏枯草可以通过提高根系APX活性, 增加叶片可溶性糖与脯氨酸的含量来清除除活性氧, 保护光系统和细胞膜的稳定性, 通过热耗散机制降低过剩光能对光合机构的破坏, 从而减少酸胁迫所造成的伤害。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|