



{kind=link}
{kind=link}
{kind=link}
{kind=link}
氮、磷添加下AMF对羊草和苜蓿生长与光合生理特性的影响
[单立文1 , 张强2 , 朱瑞芬2 , 孔晓蕾2 , 陈积山2, *  ]
]
]
|
|


作者简介:单立文(1993-),男,甘肃武威人,在读硕士。E-mail:121663667@qq.com
丛枝菌根共生对植物养分利用效率起重要作用,豆科和禾本科牧草混播可以有效提高草地产量,而适量外源氮、磷的添加同样可以促进牧草的生长。为了更好地改善对草地的利用效率,将混播、AMF以及氮、磷添加有效结合将会是其中的一条途径。因此,通过以丛枝菌根真菌的有无(+/-AMF)为主处理,不同氮、磷添加量(N0,N1,N2)为副处理,研究豆禾共存群落植株的生长及其光合作用的响应,结果表明:1)AMF对羊草和苜蓿净光合速率均有改善,而且对羊草净光合速率影响显著。2)光响应曲线分析表明,羊草和苜蓿净光合速率均随光强的增加而升高,且羊草净光合速率在+AMF条件下均高于-AMF。 3)AMF显著提高了羊草和苜蓿叶绿素含量,不同氮、磷添加量条件下,+AMF处理的羊草和苜蓿的叶绿素含量均高于-AMF处理。 4)氮、磷添加量和AMF处理显著影响羊草和苜蓿生物量,豆禾混播草地共存群落的处理以+AMF+N2处理为宜。
Symbiosis between arbuscular mycorrhizal fungi (AMF) and forage plants plays an important role in the utilization efficiency of plant nutrients in grassland. Mixed sowing of legumes and grasses can effectively increase grassland yield through biological N fixation, and the addition of appropriate amounts of external nitrogen and phosphorus can also further promote herbage growth. In order to better manage the utilization efficiency of grassland in the future, the optimal combination of plant species, AMF and nitrogen and phosphorus will need to be known. Therefore, an experiment was conducted with the presence or absence of arbuscular mycorrhizal fungi (+/- AMF) as the main treatment, and different nitrogen and phosphorus fertilizer additions (N0, no added N or P; N1, 2.0 g N and 0.2 g P·m-2; N2, 20.0 g N and 2.0 g P·m-2) as secondary treatments, to study the plant growth and photosynthesis response of coexisting alfalfa communities. It was found that: 1) AMF significantly improved the net photosynthetic rate of Leymus chinensis and Medicago sativa. 2) The light response curve indicated that the net photosynthetic rate of L. chinensis and M. sativa increased with increase in light intensity, and the net photosynthetic rate of +AMF L. chinensis was higher than -AMF L. chinensis. 3) AMF significantly increased the chlorophyll content of L. chinensis and M. sativa. Under different nitrogen and phosphorus application rates, the chlorophyll content of +AMF L. chinensis and M. sativa +AMF was higher than that of the -AMF treatments. 4) The application rate of nitrogen and phosphorus and the AMF treatment significantly affected the biomass of L. chinensis and M. sativa. In this research, treatment that optimized herbage production of coexisting communities of alfalfa-grass mixed grassland was +AMF+N2.
丛枝菌根真菌(arbuscular mycorrhizal fungi, AMF), 草地生态系统的重要组成部分, 植物群落多样性和生产力的决定因子之一[1], 可以与约80%的植物共生形成菌根, 而菌根则可以改善共生植物对土壤中氮、磷、水分以及矿质养分的吸收进而改变植物对逆境的适应生存能力[2], 提高植物抗病、抗干旱、抗盐、抗重金属等诸多特性[3, 4], 促进了植物的光合作用[5]和生长[6], 菌根还可以影响共生植物对碳水化合物的再分配, 进而间接影响群落中物种间的相互关系[7]。然而AMF对共生植物的侵染会随着外界营养状况的不同而有所变化[8]。土壤高浓度的N或P会改变根际pH值, 对菌根有一定的影响[9], 另外N
有研究表明, AMF可以与羊草(Leymus chinensis)形成良好的共生关系[11], 并且可以促进羊草幼苗磷、镁等营养元素的吸收增加叶绿素含量, 同时AMF分泌的细胞分裂素, 可增加叶绿体的发育与叶绿素水平的提高, 进而改善植株光合作用, 增加生物量并应对胁迫[12]。此外, 外源氮或者磷元素添加与AMF互作会对羊草生长产生显著影响[13, 14]。与羊草同属禾本科的玉米(Zea mays)与AMF共生后, 可以促进玉米叶片叶绿素合成, 增强菌根植株的光合能力[15], 提高玉米的净光合速率、蒸腾速率和气孔导度[16]。同时甜瓜(Cucumis melo)幼苗接种AMF后叶绿素含量、净光合速率、水分利用率、最大羧化速率均显著提高[17]。苜蓿(Medicago sativa)是一种多年生优质蛋白豆科牧草, 在盐碱地中生长会受到限制而影响其产量及推广应用, 而苜蓿与AMF共生后可以提高苜蓿P和N的含量[18], 增强苜蓿光合碳同化能力[19], 促进苜蓿生长并应对盐碱胁迫[20]。同时香椿(Toona sinensis)[21]、猕猴桃(Actinidia)[22]、柑橘(Citrus reticulata)[23]接种丛枝菌根真菌后, 植株在株高、矿质营养物质的吸收和积累、光合特性、渗透调节物质含量等方面均具有显著改善。
目前, 混播草地在我国东北地区广泛推广, 羊草和苜蓿是其栽培模式的关键草种, 其优越的草地生产性能和稳定性可超过单一的苜蓿草地或羊草草地。国内外的许多试验都表明:禾本科牧草和豆科牧草混播, 可以在稳定地提高产量的同时还提高禾本科牧草的粗蛋白质和可消化蛋白质的含量[24]。因此, 本研究通过将苜蓿和羊草混播, 研究AMF和氮、磷添加处理对混播羊草和苜蓿生长特征的影响, 提出草地建植的高效管理模式, 以期为混播草地高效利用提供理论依据, 为牧草产业良性发展提供技术支持。
羊草又名碱草, 多年生, 草甸草原的建群种, 其种子由黑龙江省农业科学院提供, 羊草自然发芽率偏低为16%, 经过处理后发芽率达40%。紫花苜蓿, 多年生豆科植物, 是改良退化羊草草地的先锋优良豆科牧草, 种子由黑龙江省农业科学院提供。试验用土取自兰西试验基地(东经125° 28'24″, 北纬46° 32'17″), 属于大陆性季风气候, 区域年降水469.7 mm, 年均气温2.9 ℃; 年积温≥ 10 ℃, 活动积温2760 ℃; 年均日照时数2713 h; 年均除霜期为9月23日, 年均终霜期为5月15日, 无霜期为130 d, 全年结冻期为183 d。土壤pH值在8.12~10.08, 土壤全磷含量为0.23 g· kg-1, 土壤全氮含量为2.15 g· kg-1。试验用盆高15 cm, 宽24 cm。
1.2.1 试验设计 本研究设置AMF(有和无)和氮磷添加量(N0, N1, N2)两个因素水平, 共6个处理, 每个处理3个重复。氮、磷添加量为相同氮、磷比例的不同量, N0:不添加N和P, N1: 添加 2.0 g N· m-2和0.2 g P· m-2, N2:添加20.0 g N· m-2和2.0 g P· m-2。AMF来源于兰西基地试验区的原位羊草草甸土, 经过鉴定其土壤主要含有3种AMF:根内球囊霉(Glomus intraradices, RI, 369个孢子每10 g)、摩西球囊霉(Glomus mosseae, FM, 282个孢子每10 g)、地表球囊霉(Glomus versiforme, GV, 310个孢子每10 g)。尿素和过磷酸钙购于黑龙江景顺农业生产资料有限公司, 其中N选择有机态尿素(CON2H4, 总氮含量≥ 46.4%), P选择有机态过磷酸钙(P2O5含量≥ 16%)。
试验用土于2018年6月中旬在原位羊草草甸草原收集010 cm土壤, 过5 mm孔径的筛去除根系石子等杂物后, 一部分土壤不经过灭菌处理作为含AMF接种剂的供试土壤, 一部分经过灭菌处理作为培养基质。依据灭菌处理的相关研究, 主要的灭菌方法有高温灭菌法、γ 射线灭菌法和化学熏蒸法。而γ 射线灭菌法在种子、土壤灭菌处理中较为常规。同时由于γ 射线采用的是低温灭菌方式, 排除了高温给土壤带来的伤害。因此, 许多学者认为, 它是土壤灭菌最有效的方式, 而且γ 射线在25 kGy时土壤灭菌效果最佳, 所以本研究选用该方法灭菌。AMF接种过程具体为, 在每盆当中分等量灭菌土壤作培养基质, 其中9盆添加过5 mm孔径的筛的新鲜土壤1000 g作为接种AMF, 而在另外9盆中加入等量灭菌土壤并添加滤液200 mL。其中滤液是将新鲜土壤与蒸馏水按体积比1∶ 1混合, 搅成泥浆后静置片刻, 用10 μ m的筛子过滤上层清液3遍, 每个盆中加入过滤液200 mL, 因为一般AMF孢子的直径大于10 μ m, 而细菌等土壤微生物较小能够通过10 μ m的筛子, 所以滤液中不含AMF孢子而含有细菌等其他土壤微生物, 培养基质中在加入该滤液后, 能够将除AMF外的其他土壤微生物补充回培养基质中, 最终保证接种和不接种AMF处理只有含AMF与否的差异。将灭菌土壤填装在专用试验装置的外层和内层, 后者的提取液按照一定量只浇散在专用试验装置的内层, 两层土壤在盆内的高度保持一致, 内外两层之间用38 μ m孔径尼龙网分隔(图1), 只有水分和AMF菌丝能够通过, 植物根系不能通过, 以消除植物根系影响。
 | 图1 专用试验培养装置Fig.1 Culture device of special test |
1.2.2 试验条件 试验于黑龙江省农业科学院温室进行, 室内平均温度26.8 ℃, 湿度42.1%, 日照时长8 h。2018年7月15进行种植, 每盆种510粒羊草和苜蓿种子, 保证每盆成活苜蓿和羊草各3株, 多的则剔除。成株后每隔4 d浇水一次使水分充足, 为保证光照均衡每隔2周进行不同区域位置互换。在进行测量前每3个月刈割一次, 留茬5 cm, 每次刈割处理之后加入等量氮、磷, 在2019年4月4开始进行光合数据和生长指标的测量。
1.3.1 生长指标的测定 2019年4月1日- 4月4日对羊草和苜蓿生长指标进行测定。分别用直尺与游标卡尺测定株高和基径。将羊草和苜蓿刈割后, 先用分析天平分别测量鲜重, 然后105 ℃杀青0.5 h, 最后65 ℃烘72 h, 称量其干重。
1.3.2 光合生理指标的测定 采用Li-6400便携式光合作用测定系统(LI-COR, Li-6400XT, 美国) 测定光合特性和光响应, 每个处理选取3株, 每株取1片完整叶, 测定结果取平均值。测定参数包括羊草和苜蓿叶片的净光合速率(photosynthetic rate, Pn)、气孔导度(stomatal conductance, Gs)、胞间CO2浓度(intercellular CO2 concentration, Ci)和蒸腾速率(transpiration rate, Tr)。
在上午9:00-12:00测定光响应曲线, 采用LED光源控制光合有效辐射, 其光合有效辐射依次为1000, 750, 500, 150, 60, 0 μ mol· m-2· s-1。光响应曲线模型用直角双曲线修正模型, 同时用SPSS软件对光响应曲线进行拟合, 得到表观量子效率(apparent quantum efficiency, A), 最大净光合速率(maximum net photosynthetic rate, Pnmax), 光补偿点(light compensation point, LCP), 光饱和点(light saturation point, LSP)和暗呼吸速率(dark breathing rate, Rd)。
1.3.3 叶绿素相对含量测定 采用CCM-300手持式叶绿素测定仪(OPTI-SCIENCES, 美国)测定叶绿素相对含量。
1.3.4 羊草和苜蓿侵染率的测定 将所取的羊草和苜蓿根, 剪切成23 cm后分别装入标记的10 mL离心管, 加入适量FAA(formaldehyde-acetic acid-ethanol fixative)固定液保存, 然后使用曲利苯蓝染色液染色, 经过脱色后制片并在电镜下观察统计, 最后使用Mycocalc.exe软件统计分析侵染率。
采用SPSS 17.0软件进行双因素方差分析(ANOVA), 采用SigmaPlot 12.0绘图。
中午12:00时光照相对较强, 各处理条件下AMF均提高了羊草净光合速率和光能利用率, 降低了胞间二氧化碳。而且在N1时, AMF对羊草净光合速率影响显著, 同时在N1和N2时, AMF对羊草光能利用率影响显著(表1)。胞间二氧化碳与净光合速率呈负相关, AMF降低了胞间二氧化碳, 且在N0和N1时, AMF对胞间二氧化碳影响显著。同时氮、磷添加量(N0, N1, N2)对羊草各光合特性参数也有影响, 其中气孔导度在+AMF+N2时比+AMF+N1低, 且两种处理间差异显著, -AMF+N2比-AMF+N1低且差异显著。受氮、磷添加量的影响蒸腾速率在+AMF和-AMF条件下, N2相对N1下降显著。双因素方差分析表明, +AMF显著影响羊草净光合速率、胞间二氧化碳、光能利用率, 而氮、磷添加量对气孔导度和蒸腾速率有显著影响, 但AMF和氮、磷的交互作用对各处理影响均不显著(表1)。
| 表1 氮、磷添加条件下有无丛枝菌根真菌对羊草幼苗叶片光合特性参数的影响 Table 1 Effects of arbuscular mycorrhizal fungi on photosynthetic characteristics of L. chinensis seedlings under nitrogen and phosphorus addition |
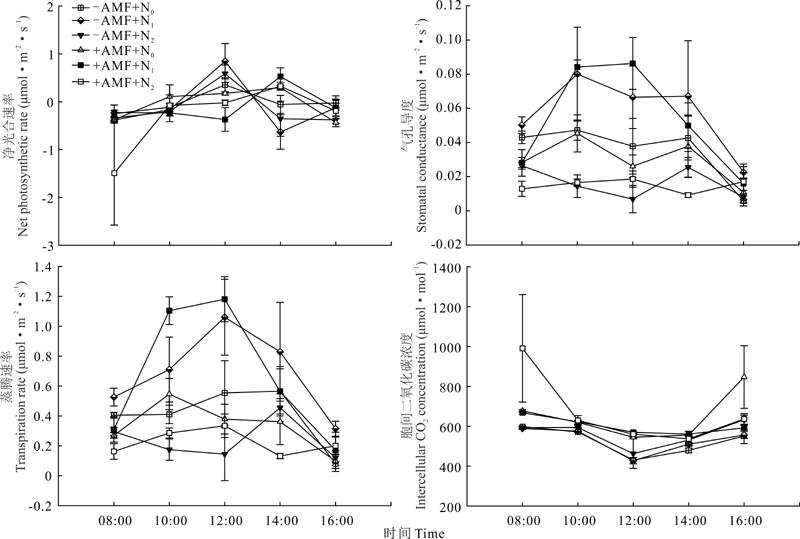 | 图2 氮、磷添加与接种丛枝菌根真菌对羊草幼苗叶片光合特性日变化的影响Fig.2 Effects of nitrogen and phosphorus addition and inoculation with arbuscular mycorrhizal fungi on diurnal changes of photosynthetic characteristics of L. chinensis seedlings |
光合日进程中, 羊草净光合速率在-AMF时增加明显, 在12:00时达到峰值, 然后出现下降趋势, 14:00呈现光合午休现象。+AMF时净光合速率增加缓慢, 在14:00才达到峰值, 然后开始逐渐下降。随着日照和温度改变, 各处理下蒸腾速率均升高, 在12:00时+AMF+N1蒸腾速率达最高, 且与+AMF+N0和+AMF+N2间差异明显。整体胞间二氧化碳与净光合速率的变化呈现相反的趋势(图2)。
同理, 随着光照变化, 从8:00-16:00苜蓿净光合速率在+AMF+N0处理下, 分别在10:00和14:00 呈现明显的双峰现象(图3)。胞间二氧化碳和净光合速率变化呈相反的趋势。
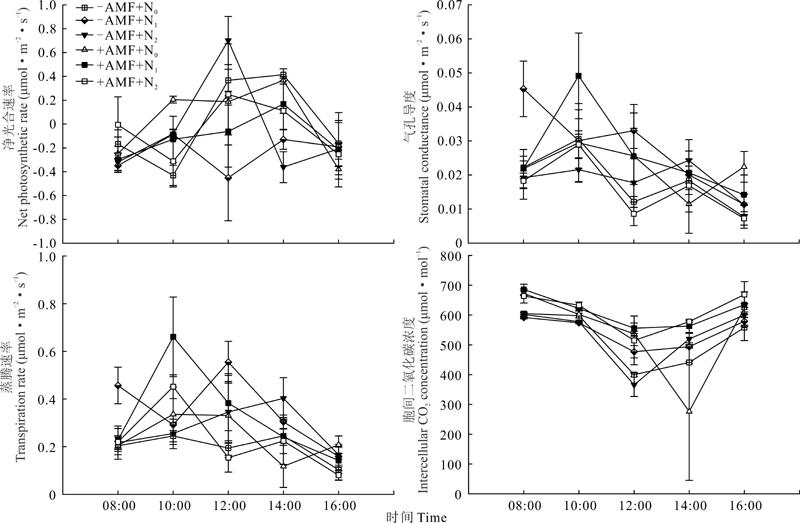 | 图3 氮、磷添加条件下有无丛枝菌根真菌对苜蓿幼苗叶片光合特性日变化的影响Fig.3 Effect of arbuscular mycorrhizal fungi on the diurnal changes of photosynthetic characteristics of M. sativa seedlings under nitrogen and phosphorus addition conditions |
苜蓿净光合速率受AMF影响, 在+AMF+N0和+AMF+N2分别比-AMF+N0和-AMF+N2时均有提高, 但差异并不显著。AMF对气孔导度、蒸腾速率、水分利用率、光能利用率的影响也不显著, 但胞间二氧化碳在+AMF+N0和+AMF+N2分别比-AMF-N0和-AMF-N2低, 而且差异显著。苜蓿净光合速率随着氮、磷添加量的增加出现先降低后升高现象, 而且在+AMF+N2与+AMF+N1间差异显著, 苜蓿其他光合特性参数均受氮、磷影响不显著(表2)。双因素方差分析表明AMF除对苜蓿胞间二氧化碳影响显著外, 对其他各参数影响均不显著。氮、磷和AMF与氮、磷互作对苜蓿各参数影响均不显著(表2)。
| 表2 氮、磷添加条件下有无丛枝菌根真菌对苜蓿幼苗叶片光合特性参数的影响 Table 2 Effects of arbuscular mycorrhizal fungi on photosynthetic characteristics of M. sativa seedling leaves under nitrogen and phosphorus addition conditions |
羊草和苜蓿叶片净光合速率在不同处理下均随光合有效辐射增强而升高并在达最大值后开始趋于稳定。当光合有效辐射为500 μ mol· m-2· s-1时, 羊草净光合速率达最大, 且在N0, N1, N2时+AMF羊草的净光合速率均高于-AMF, 其中在+AMF+N1 条件下羊草净光合速率最高。在光合有效辐射为750 μ mol· m-2· s-1时, 苜蓿净光合速率达到最大, 表现为有无AMF条件下N0处理的苜蓿净光合速率均高于N1和N2处理(图4)。
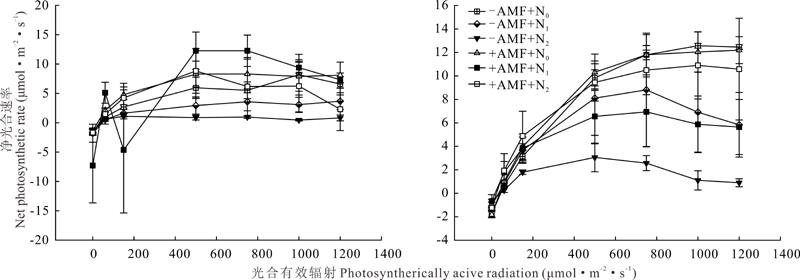 | 图4 氮、磷添加与有无AMF对羊草(左)和苜蓿(右)叶片光响应的影响Fig.4 Effect of nitrogen and phosphorus addition and presence or absence of AMF on light response of L. chinensis (left) and M. sativa (right) leaves |
直角双曲线修正模型拟合(表3, 4)结果表明, 相同氮、磷添加量下, +AMF的最大净光合速率均高于-AMF的最大净光合速率。+AMF和-AMF时, 羊草最大净光合速率随氮、磷添加量的增加呈下降趋势(表3)。方差分析进一步显示, AMF和N/P互作对羊草光补偿点影响显著(表3)。
| 表3 氮、磷添加与有无丛枝菌根真菌对羊草幼苗叶片光合特性参数的影响 Table 3 Effects of arbuscular mycorrhizal fungi on photosynthetic characteristics of L. chinensis seedlings under nitrogen and phosphorus addition conditions |
接种AMF提高苜蓿最大净光合速率, 在N0, N1, N2条件下+AMF苜蓿最大净光合速率均高于-AMF处理, 而且苜蓿最大净光合速率在+AMF+N0处理时比-AMF+N0增加30.46%, 差异显著(表4)。方差分析表明, 苜蓿最大净光合速率受氮磷添加影响显著, 苜蓿光饱和点受AMF和N/P互作影响显著, 而表观量子效率、光补偿点、暗呼吸速率均无显著变化。
| 表4 氮、磷添加与有无丛枝菌根真菌对苜蓿叶片光合特性参数的影响 Table 4 Effects of arbuscular mycorrhizal fungi on photosynthetic characteristics of M. sativa seedlings under nitrogen and phosphorus addition conditions |
AMF显著影响羊草叶绿素含量。随着氮、磷添加量的增加, 羊草叶绿素含量表现出先增加后降低的趋势, 且差异显著(表5)。方差分析表明, +/-AMF、N/P、AMF和N/P互作均对羊草和苜蓿叶绿素含量产生显著影响。
| 表5 氮、磷添加与有无AMF对羊草和苜蓿叶绿素含量的影响 Table 5 Effect of inoculation of AMF on the content of chlorophyll in L. chinensis and M. sativa under the conditions of nitrogen and phosphorus addition (mg· m-2) |
AMF促进了羊草的生长, 对羊草基茎、茎鲜重、叶鲜重、总鲜重均有改善, 但只在总鲜重方面影响显著(表6)。氮、磷添加促进羊草生长, +AMF+N2处理与+AMF+N0处理相比, 羊草株高和总鲜重分别增加了15.5%和107.9%。方差分析显示, 氮、磷添加对羊草总鲜重有显著影响。
| 表6 氮、磷添加与有无AMF对羊草生长的影响 Table 6 Effect of inoculation of AMF on the growth of L. chinensis under the conditions of nitrogen and phosphorus addition |
氮、磷添加量处理显著提高了苜蓿茎鲜重和叶鲜重, +AMF条件下N1和N2处理的苜蓿茎鲜重分别比N0处理增加87.5%和130%, -AMF条件下N1和N2处理分别比N0处理增加109%和174%, 叶鲜重在+AMF条件下N1和N2处理分别比N0处理增加102%和172%(表7)。AMF增加了苜蓿株高, 在N0时+AMF相对-AMF株高增加73.65%, 差异显著。在N1和N2时+AMF总鲜重显著高于-AMF, 且受氮磷添加量的增加苜蓿鲜重也显著增加。方差分析结果显示, N/P添加对苜蓿茎鲜重和叶鲜重有显著影响, AMF对苜蓿株高影响显著(表7)。
| 表7 氮、磷添加与有无AMF对苜蓿生长的影响 Table 7 Effect of inoculation of AMF on the growth of M. sativa plant seedlings under the conditions of nitrogen and phosphorus addition |
氮、磷处理对羊草和苜蓿AMF的侵染率有影响, 但对苜蓿影响并不显著。+AMF处理结果中羊草在N2时菌根侵染率达最大, 与N0, N1间差异显著(表8)。 -AMF处理结果中未观察到对羊草和苜蓿根系有侵染。
| 表8 氮、磷添加对羊草和苜蓿AM菌根侵染率的影响 Table 8 Effect of nitrogen and phosphorus addition on AM infection rate of L. chinensis and M. sativa (%) |
AMF与植物共生的现象比较普遍, 但不同的植物对菌根的依赖程度各有差异。本研究中选用的羊草为C3型多年生草本植物, 紫花苜蓿为多年生豆科植物, 结果显示其根系均能够与AMF形成良好的共生关系, 这与甄丽娜等[13, 14]和才华等[25]的研究结果一致。此外, AMF对植物的依赖程度还受温度[26]、根际土壤氮、磷含量的影响, 在本研究结果中也证实了这一点。羊草和苜蓿侵染率受氮磷添加升高, 均在+AMF+N2时达到最高, 这可能是由于本研究中氮、磷添加量未超出菌根共生的极限范围, 而在该范围内有利于菌根真菌的共生, 当然也有相关试验表明, 氮、磷含量过高时会抑制AMF侵染, 这可能是高浓度的N或者P会改变根际pH值对菌根产生影响[9], 而NH4+的大量存在也可能对AMF有毒害作用。
AMF与植株共生后增加了根吸收面积, 还可以进入植株无法抵达的土壤孔隙吸收水分和养分, 促进了植物根系对矿质营养和水分的吸收[27], 因此可以改善宿主植物生长。然而, 国内外对AMF的研究多集中体现在生物量、氮磷含量等方面, 而对共生植株生长过程特性和作用机制的研究相对较少。研究发现, N和P元素与植物光合作用密切相关, P元素是植物光合作用所需要的酶以及ATP的重要组成元素, 磷含量的增加有利于植株光合作用进行[26], 而N元素则可以对光合电子传递链的相关的酶活性和含量进行调控[28], 且在一定范围内植物氮素含量的提高将促进光合作用[29], 因此 AMF改善共生植株的生长与植株光合作用密切相关, 本研究结果与此一致:AMF对羊草和苜蓿净光合速率均有改善, 在N1时AMF对羊草净光合速率影响显著。同时郑舜怡等[30]和Wang等[31]的研究也表明, AMF能显著提高辣椒(Capsicum annuum)和生姜(Zingiber officinace)植株叶片净光合速率、蒸腾速率和气孔导度。这可能是AMF增强氮、磷相关代谢酶活性促进植物氮、磷元素的吸收, 一方面AMF侵染植物后菌丝分泌有机酸和磷酸酶, 利于土壤中难溶性P被植株利用[32, 33]。苏友波等[34]在对玉米接种AMF后结果表明AMF对根际土壤磷酸酶活性有增强作用。另一方面氮素的转化主要受脲酶、硝酸还原酶等的影响, 而AMF则可以增强该类相关酶活性而促进共生植物对氮素的利用[35]。
植物生长与光合作用密切相关, 而光合作用受限于叶绿素和光强等因素, 因此植物生长与叶绿素和光强紧密相关。本研究结果发现, AMF显著提高了苜蓿和羊草叶绿素含量, 这可能是由于接种AMF的植株一方面能促进镁等营养元素的吸收进而增加叶绿素含量, 另一方面AMF分泌物质如细胞分裂素, 可增加叶绿体发育与提高叶绿素水平[36]。同时本研究结果还表明, AMF和氮、磷添加交互作用对羊草和苜蓿叶绿素含量影响显著, 可能是由于AMF侵染植株后促进了添加氮的吸收, 而氮是叶绿素的主要构成元素, 周霞等[37]的研究结果也发现, 接种AMF会提高植物叶绿素含量。相反, 添加的氮、磷则可以通过影响AMF的侵染间接影响植株叶绿素含量。例如, 甄丽娜等[13, 14]对羊草接种AMF后发现, AMF侵染率受氮或磷浓度影响。光强受限时光强的增加使植物净光合速率升高, 本研究结果表明羊草在500 μ mol· m-2· s-1时净光合速率达最大, 而苜蓿则在750 μ mol· m-2· s-1时净光合速率达最大, 通过对光响应结果拟合发现羊草和苜蓿最大净光合速率在+AMF时均高于-AMF处理, 这可能是AMF增强了植物对光能的利用[37]。
研究发现氮、磷含量对植株生长的影响与所处环境的氮、磷含量相关[38, 39], 当植株生长受氮、磷限制时, 添加氮磷量会促进植株生长[40], 而当植株所处环境氮、磷含量达到饱和时植株生长会随氮、磷添加量的增加而受抑制[41]。然而, 在本研究结果中, 羊草和苜蓿总生物量随着氮、磷水平的增加而增加, 而且接种AMF处理均高于未接菌处理, 这可能是由于本研究所用根际土壤氮、磷含量有限, 而低磷土壤中AMF对植株生长有促进作用[42]。此外, 有研究表明羊草和苜蓿混播后有利于草地产量增加[43]。这是由于豆科牧草与羊草混播时, 羊草不断吸收和利用土壤有效氮使土壤氮含量至低水平, 从而间接刺激豆科牧草的固氮能力[44]。而接种AMF后的混播羊草和苜蓿又可以通过地下连接不同植物根系的丛枝菌根的菌丝将氮素直接传递给羊草进一步增强苜蓿固氮能力[45]。
研究发现, 不同处理条件下羊草和苜蓿光合日进程均有峰值出现。AMF显著提高混播羊草净光合速率, 同时对苜蓿净光合速率也有改善, 但影响并不显著。光响应曲线分析表明, 羊草和苜蓿净光合速率均随光强的增加而升高, 羊草净光合速率在+AMF条件下均高于-AMF。AMF显著提高了羊草和苜蓿叶绿素含量, 不同氮、磷添加量条件下, +AMF处理的羊草和苜蓿的叶绿素含量均高于-AMF处理。氮、磷添加量和AMF处理显著影响羊草和苜蓿生物量, 豆禾混播草地共存群落的处理条件以+AMF+N2为宜。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|