

{kind=link}
{kind=link}
干热河谷冲沟沟头活跃度对植物群落结构的影响
[何周窈1, 2  , 王勇
, 王勇1, * , 苏正安2 , 杨鸿琨3 , 周涛2 ]
, 王勇, 苏正安|
|


作者简介:何周窈(1994-),女,四川雅安人,在读硕士。 E-mail: thezhouyaom@163.com
为研究干热河谷冲沟沟头活跃程度对植被物种组成及其群落结构的影响,从而服务于冲沟植被恢复过程中最佳植被配置,采用典型样地法对元谋县内22条冲沟进行实地调查。研究结果表明:1)冲沟内植物种类随着沟头的稳定程度增加而增加,其中活跃型冲沟内植物12科28属共28种,半稳定型冲沟内植物16科29属共29种,稳定型冲沟内植物15科35属共37种;2)不同活跃程度的冲沟植被群落按生活型可分为3个层次,一是以扭黄茅为优势种,孔颖草、三芒草为次优种的草本层;二是以车桑子为优势种,银合欢为次优种的灌木层;三是以印楝为优势种,桉树为次优种的乔木层,但活跃型冲沟无乔木层;3)冲沟乔木层、灌木层和草本层的植被盖度均随着冲沟沟头稳定性的增加而增加,表现为稳定型冲沟>半稳定型冲沟>活跃型冲沟;4)不同活跃程度的冲沟沟头群落结构均以多年生草本和灌木为主,乔木种类较少,优势物种相同且占有显著地位,但随着冲沟沟头的逐渐稳定,乔木数量和多样性均逐渐增加;5)活跃型冲沟、半稳定型冲沟、稳定型冲沟之间草本层、灌木层、乔木层的植物群落相似性分别表现为中等相似、极为相似、极度不相似,且冲沟不同恢复阶段灌木层中植物有趋同演化的趋势,共有程度很高,冲沟沟头活跃程度对灌木层植物群落功能造成的影响小于对草本层和乔木层植物群落。总体而言,不同活跃程度的冲沟沟头植被群落结构之间具有显著差异,研究结果将为冲沟生态恢复提供科学依据。
In this research, field surveys of species composition and vegetation community structure of 22 gullies in the hot-dry valley region of Yuanmou County in Yunnan Province were carried out using standard vegetation sampling methods to study the plant community structure in gullies with different activity levels. As the gully stability increased, the number of plant species also increased. Twenty-eight species of plants belonging to 28 genera of 12 families, were found in active gullies. Twenty-nine species of plants belonging to 29 genera of 16 families, were found in semi-stable gullies. Thirty-seven species of plants belonging to 35 genera of 15 families were discovered in stable gullies. Based on plant form, the gully vegetation communities could be divided into three layers (herb layer, shrub layer and tree layer) which varied with the activity level of the gullies. No tree layer was found in the active gullies. The dominant species in the herb layer was Heteropogon contortus, with Bothriochloa pertusa and Aristida adscensionis as sub-dominant species. The dominant species in the shrub layer was Dodonaea viscosa, with Leucaena leucocephala as the sub-dominant species. The dominant species of the tree layer was Azadirachta Indica, with Eucalyptus robusta as the sub-dominant species. As the gully stability increased, so also did the coverage of its vegetation community. The plant communities in gullies with differing activity levels were all dominated by perennial herbs and shrubs, with a smaller number of tree species of trees, in which the dominant species were the same and played similar roles in the plant communities. However, with increasing stability of gullies, the number and diversity of trees gradually increased. For active gullies, semi-stable gullies and stable gullies, the plant community similarities were comparatively similar in the herb layer, extremely similar in the shrub layer, and extremely different in the tree layer. As the stability of a gully changed, the tendency for the plant community matured without species change in the shrub layer was evident. However, for the herb layer and tree layer, as the gully stability increased with time the species composition of the vegetation community evolved. These results indicate that the activity status of a gully plays an important role in determining the community structure of the tree layers, while the community structure of the shrub layer was less affected by the activity status of the gully. In summary, an obvious difference in vegetation structure was evident between gullies with different activity status, and this information could provide a scientific basis for gully ecological restoration decisions and for formulation of future studies.
在生态脆弱区和土壤严重退化区, 植被恢复是生态修复的主要措施[1, 2, 3]。目前, 全世界面临着土壤退化和耕地资源减少的巨大压力。据估测, 世界中度及以上退化程度的土壤面积约13.52× 108 hm2, 亚洲约4.91× 108 hm2 , 生态修复问题亟待解决[4]。
金沙江干热河谷是我国西南地区典型的生态脆弱区和土壤严重退化区。80 年代末期该地区森林覆盖率仅为 5.2%, 土壤侵蚀模数达2547 t· km-2· a-1[5], 在元谋县境内, 土林总面积达50 km2, 盆地内沟壑密度为3.0~5.0 km· km-2, 年均溯源侵蚀速度50 cm 左右, 土壤侵蚀模数高达1.64 万 t· km-2· a-1[6]。经过多年生态修复, 元谋县森林覆盖率已经达到43.1%, 大量冲沟发育区植被已经开始逐渐恢复。但由于前期该区土壤侵蚀严重、如今生态恢复仍面临较大困难, 尤其是在冲沟沟头, 由于沟头溯源侵蚀强烈, 不断蚕食农地、损坏公路等, 给当地的经济发展带来了巨大阻碍。对此, 前人在冲沟沟头的水土流失机理[7, 8]、冲沟侵蚀与预报[9]、适宜植被类型及水保效益[5, 10, 11]、人工建林技术路线[12]、不同恢复措施的植被群落多样性[13]、生态修复及评价[14, 15]等方面开展了大量研究, 提出了一系列的治理模式防治该区的水土流失问题, 如:重度以上退化生态系统推广混交模式与自然禁封治理模式, 混交类型以灌草为主, 条件较好的地段可进行乔灌草行间混交, 多林草注意豆科和非豆科的搭配等[5, 12, 16]。但以上研究的时间尺度均较短, 且多以水分、坡度为主要考虑因素, 且该区冲沟活跃程度不同, 土壤退化程度也不尽相同, 进行大面积冲沟植被恢复时常因冲沟活跃程度等差异较大导致立地条件出现显著差异, 恢复效果欠佳[17]。与此同时, 针对不同活跃程度的冲沟沟头植被群落结构特征的研究也较少。因此, 干热河谷冲沟发育区的植被恢复需结合冲沟沟头的活跃程度, 制定适宜于不同冲沟沟头活跃程度的植被恢复措施, 以避免造成人力、物力和财力的巨大浪费。
本研究选择金沙江干热河谷区的典型区域— 元谋县, 调查该区不同活跃程度的冲沟沟头植被现状, 并运用数量生态学和恢复生态学理论, 结合沟头的不同活跃程度, 对冲沟植物群落的盖度、重要值、多样性指数及相似性等指标进行分析, 阐明冲沟沟头活跃程度对植物群落结构特征的影响, 探索不同活跃程度的冲沟沟头的最优植被配置, 从而为该区域的植被恢复策略制定提供理论依据。
本研究区位于云南省楚雄彝族自治州元谋县(101° 35'-102° 06' E, 25° 23'-26° 06' N), 属于典型的金沙江干热河谷区, 生境炎热干燥, 植被类型类似于热带稀树灌丛草原[6]。境内最高海拔2835.9 m, 最低海拔898 m, 年均气温21.9 ℃, 年日照时数2670.4 h, 多年平均降水量626 mm, 年均潜在蒸发量3911 mm, 约为降水量的6倍[6]。金沙江干热河谷区土壤以燥红土和变性土为主。其中变性土土壤肥力很差, 土面裂隙多, 土质粘重, 膨胀收缩性强; 燥红土是该区的基带土壤, 土壤砂粒含量较高, 表层往往覆盖一层铁锰胶膜, 入渗性较差[18]。金沙江干热河谷区的地带性植被为亚热带西部半湿润常绿阔叶林和针叶林, 大面积分布灌丛草被, 较小面积分布了合欢(Albizia julibrissin)、人工桉树(Eucalyptus robusta)林等乔木。该区乡土草被以扭黄茅(Heteropogon contortus)、旱茅(Ere-mopogon delevayi)、孔颖草(Bothriochloa pertusa)等多年草本植物为主, 乔木以余甘子(Phyllanthus emblica)、酸角(Tamarindus indica)、山合欢(Albizzia kalkora)、木棉(Bombax ceiba)为主, 灌木以车桑子(Dodonaea viscose)为主。总体而言, 该区灌、乔木稀疏分布, 草被为主, 植被覆盖度较低[19]。
试验区位于中国科学院、水利部成都山地灾害与环境研究所与云南省农科院共建的元谋干热河谷沟蚀崩塌试验站。该试验站地处金沙江干热河谷冲沟发育区内, 且冲沟保存完整, 具有典型性和代表性。在研究区选定22条人为活动影响较弱的典型封禁冲沟, 依据冲沟沟头的活跃程度和形态划分为活跃型冲沟、半稳定型冲沟和稳定型冲沟3种类型, 其中活跃型冲沟9条, 半稳定型冲沟5条, 稳定型冲沟8条。活跃沟头沟壁陡立, 在水力和重力侵蚀作用下, 往往呈“ 上凸下凹” 的形状, 每年均伴有沟头延伸、沟壁发育, 跌坎明显, 常见新鲜崩积物[16, 20]; 半稳定型冲沟的沟头跌坎较低, 沟壁扩张速度缓慢或停止扩张, 偶见新鲜崩积物; 稳定型冲沟的沟头无明显跌坎, 沟头停止前进, 沟底宽且平坦, 无崩塌发生[20]。
样方调查时间为2018年4月20―30日。由于冲沟发育区地形破碎, 千沟万壑, 沟头面积大小不一, 故选择主观取样法。在每条冲沟的集水区及沟头部位分别设置乔木样地面积为20 m× 20 m, 灌木样地面积为5 m× 5 m, 草本样地面积为1 m× 1 m, 共设调查样方110个, 对面积小于10 m2的则全部调查。调查及测定方法参考《陆地生物群落调查观测与分析》[21]。 在所设样方中记录植物种类、高度、盖度、胸径和频度共5个指标。用于计算群落重要值、多样性指数及相似性。
| 表1 冲沟基本情况一览表 Table 1 List of basic conditions of gullies |
调查数据采用Microsoft office Excel 2010 软件进行整理, 采用SPSS 20.0软件进行统计分析, 使用Origin 8进行绘图。
1.3.1 重要值和多样性指数 根据统计的样地资料计算群落各植物物种相对频度(relative frequency, RF)、相对盖度(relative cover, RC)、相对密度(relative density, RD)、相对高度(relative height, RH)、相对显著度(relative prominence, RP)、群落物种重要值(important value, IV)。结合各层次植物的生长习性和生长情况, 所用计算公式为[21, 22]:
乔木重要值:IV=(RF+RD+RP)/3
灌木和草本重要值:IV=(RF+RC+RH)/3
基于物种重要值计算植物Simpson多样性指数(Simpson diversity index, D):
Shannon-Wiener 多样性指数(Shannon-Wiener diversity index, H):
Pielou 均匀度指数(Pielou’ s evenness index, E):E=H/lnS
修正的Hill 指数(improved Hill index, E'):E'=(
式中:S为群落中植物种数, Pi为种i重要值占所有种重要值之和的比例[21, 22]。e为常数, 等于2.718。
1.3.2 群落相似性 采用Sorensen相似性系数(similarity coefficient, SC), 对研究区域内活跃、半稳定、稳定3种类型的冲沟中的植物群落的相似性进行分析, 计算公式[21, 22]:
SC=2ω /(a+b)
式中:SC表示植物群落相似性系数; ω 表示两个植物群落中共有物种; a表示第1个植物群落中物种总数; b表示第2个植物群落中物种总数。根据群落相似性系数SC数值大小, 可划分为4个区间表示不同的相似程度:0.00~0.25、0.25~0.50、0.50~0.75、0.75~1.00分别代表极不相似、中等不相似、中等相似、极为相似。
调查发现, 不同活跃度的冲沟沟头植被群落大致可分为3个层次:一是以扭黄茅、孔颖草、三芒草(Aristida adscensionis)等禾本科植物为主的草本层; 二是以车桑子、银合欢(Leucaena leucocephala)为主的灌木层; 三是以印楝(Azadirachta indica)为主的乔木层。草本层植被盖度在活跃型冲沟中为39.96%, 半稳定型冲沟为79.20%, 稳定型冲沟为86.32%; 灌木层植被盖度在活跃型冲沟中为22.73%, 半稳定型冲沟为30.00%, 稳定型冲沟为47.52%; 乔木层植被盖度在稳定型冲沟中为5%, 半稳定型冲沟均低于5%, 活跃型冲沟无乔木(图1)。由此可见, 不同层次植物群落的植被盖度均随着冲沟沟头稳定性的增加而增加。
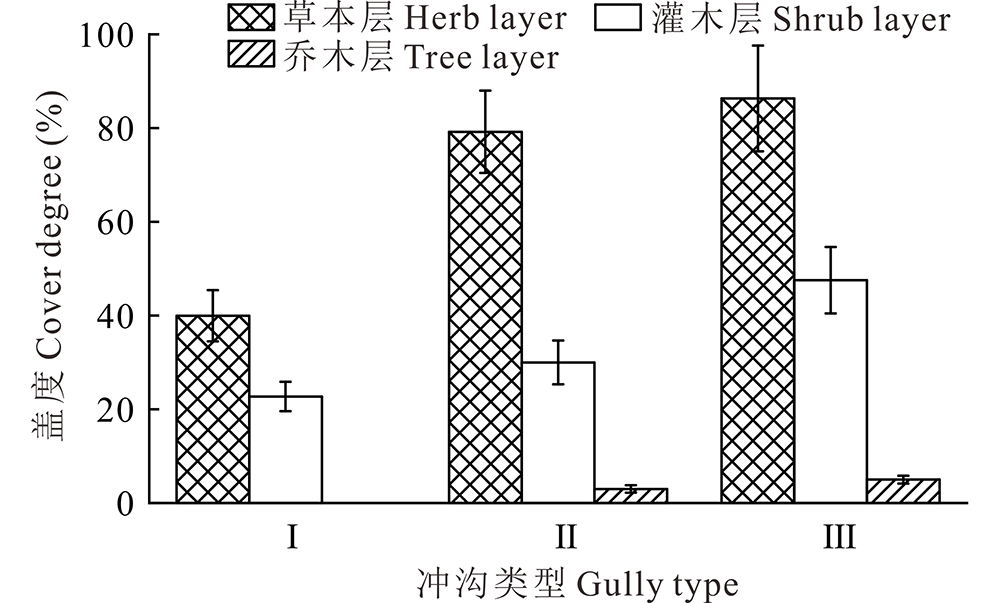 | 图1 不同活跃程度的冲沟各层植物盖度 Ⅰ .活跃型冲沟Active gully; Ⅱ .半稳定型冲沟Semi-stable gully; Ⅲ .稳定型冲沟Stable gully.Fig.1 The vegetation coverage of each layer of gully with different activity degree |
除乔木层外, 不同活跃程度的冲沟沟头植物群落拥有相同的优势种。草本层优势种为扭黄茅, 其盖度分别为:活跃型冲沟23.28%、半稳定型冲沟35.56%、稳定型冲沟52.20%; 而灌木层优势种是车桑子, 其盖度分别为:活跃型冲沟为4.40%、半稳定型冲沟15.00%、稳定型冲沟18.52%(表2)。由此可见, 冲沟沟头的活跃程度对扭黄茅和车桑子的影响小于其他植被, 可在冲沟治理工程中将这两种植被作为先锋物种进行种植。
| 表2 冲沟植被类型、盖度及重要值 Table 2 The vegetation composition and its important value of the different gully |
2.2.1 不同活跃程度冲沟沟头物种组成 研究区内植物共有48种, 分属于20科46属, 建群种7种, 广布种34种, 样地内或样地外偶见种7种(表2)。
不同活跃程度的冲沟沟头, 所拥有的植物种类和类型均不相同。活跃型冲沟内植物共28种, 分属于12科28属, 以禾本科(7属7种)、蝶形花科(6属6种)和菊科(4属4种)为主, 分别占总物种数的25.00%、21.43%、14.29%; 其余的植被分属于9科11属; 大戟科、豆科均2属2种, 共占14.29%; 龙舌兰科、鼠李科、爵床科、锦葵科、无患子科、椴树科、萝藦科均1属1种, 共占25.00%。半稳定型冲沟内植物共29种, 分属于16科29属, 以禾本科(5属5种)和蝶形花科(6属6种)为主, 分别占总物种数的17.24%、20.69%; 其余植物分属于11科18属; 其中菊科、大戟科、豆科、锦葵科均2属2种, 共占27.59%; 茄科、葡萄科、龙舌兰科、旋花科、松科、楝科、鼠李科、爵床科、无患子科、椴树科均1属1种, 共占34.48%。稳定型冲沟内植物共37种, 分属于15科35属, 以禾本科(10属10种)、蝶形花科(6属6种)和菊科(5属5种)为主, 分别占总物种数的27.03%、16.22%、13.51%; 其余16种分属于12科14属; 大戟科、豆科、锦葵科均2属2种, 共占16.22%; 桑科1属2种, 占5.41%; 桃金娘科、木棉科、龙舌兰科、楝科、鼠李科、爵床科、无患子科、椴树科1属1种, 共占21.62%。可见随着冲沟沟头稳定性的增加, 植被种类愈加丰富, 并且均以禾本科、蝶形花科、菊科等适应性强、耐旱、丛生植物为主。
2.2.2 不同活跃程度冲沟沟头植物群落重要值 对比所有植物的重要值可以发现, 不同活跃程度的冲沟沟头、不同植物群落的优势种相同, 且均在群落中拥有明显高于其他伴生种的重要地位。草本层优势种为扭黄茅, 其重要值分别为:活跃型冲沟39.87%、半稳定型冲沟28.19%、稳定型冲沟37.08%; 灌木层的优势种均为车桑子, 其重要值分别为:活跃型冲沟22.19%、半稳定型冲沟34.87%、稳定型冲沟27.86%; 乔木层中, 除活跃型冲沟无乔木外, 半稳定型冲沟和稳定型冲沟的优势种均为印楝, 其重要值分别为84.32%、21.10%。对比所有植被群落的重要值可以发现:活跃型冲沟、半稳定型冲沟、稳定型冲沟草本层和灌木层优势种的重要值比较接近, 均在90.00%以上; 其中草本层的生活型功能群均以多年生植物为优势类群, 其重要值依次是:活跃型冲沟87.05%、半稳定型冲沟72.64%、稳定型冲沟79.49%; 为一年生的植物重要值的6.73、2.65、3.99倍。乔木层的重要值差异较小, 稳定型冲沟是半稳定型冲沟的1.26倍。由此可见, 不同活跃程度的冲沟其植被结构均以多年生草本搭配灌木为主, 随着冲沟沟头稳定性的增加, 草本层和灌木层依旧占据重要地位, 而群落中原本处于劣势地位的乔木层的重要性逐渐提升, 群落结构也逐渐完善。
2.2.3 不同活跃程度冲沟沟头植物群落多样性 基于重要值对不同活跃程度的冲沟沟头、不同层次植物群落的多样性指数等进行计算(表3), 发现不同活跃程度的冲沟沟头植物群落Simpson指数和Shannon-Wiener指数均差异性显著, 且大多数呈现出稳定型冲沟> 半稳定型冲沟> 活跃型冲沟的趋势(P< 0.05); 而在不同群落类型之间, 冲沟植物群落Simpson指数和Shannon-Wiener指数大多数表现为:草本> 灌木> 乔木, 其中草本层和乔木层的植物多样性均随冲沟沟头稳定性的增加而呈现出逐渐增大的趋势, 而灌木层植物的多样性指数的最大值则出现在半稳定型冲沟。不同植被群落多样性指数的变化表明群落的多样性与冲沟沟头的活跃程度密切相关。随着冲沟沟头逐渐趋于稳定, 提供给各植物群落的生长环境越来越有利, 其草本层和灌木层植被多样性呈增长趋势, 灌木层植被多样性虽有所下降, 但仍处于较高水平。
| 表3 不同活跃程度冲沟群落植被群落特征分析 Table 3 Analysis on characteristics of gully vegetation community |
冲沟植物均匀度变化情况有所不同(表3), 不同活跃程度的冲沟沟头之间灌木的Pielou指数差异较小, 而草本与乔木的Pielou指数则存在较大差异。其中各冲沟灌木层的Pielou指数几乎无变化(P> 0.05), 而草本层和乔木层的植物Pielou指数差异性显著(P< 0.05), 且均随着冲沟沟头稳定性的增加而逐渐增加。不同活跃程度的冲沟沟头之间, 灌木层植物修正的Hill指数随着冲沟稳定性的增加而逐渐增加, 但差异性并不显著; 而草本层和乔木层植物修正的Hill指数随着冲沟稳定性的增加而逐渐减小, 且各冲沟间的修正Hill指数差异显著。结果表明活跃型冲沟的各层植被存在优势种密度较稳定型冲沟大的情况, 占据了较多的生长空间, 更易形成单一植被群落, 不利于植被群落的稳定。
2.2.4 不同活跃程度冲沟沟头植物群落相似性 各类型冲沟沟头的群落相似性分析结果显示(表4), 冲沟沟头的稳定性与植物群落的相似性密切相关, 冲沟沟头间稳定性差异越大, 植物群落相似程度越低。冲沟植物群落相似性系数大小依次为:活跃型冲沟与半稳定型冲沟(0.77)> 半稳定型冲沟与稳定型冲沟(0.70)> 活跃型冲沟与稳定型冲沟(0.65)。此外, 各阶段冲沟不同植物层之间的群落相似程度也存在明显差异(图2), 灌木层植物群落相似性最高, 表现为极度相似; 乔木层植物群落的相似性最低, 表现为极度不相似; 草本层植物群落相似性系数则表现为中等相似。由此可见, 冲沟灌木层植物群落之间的相似性均大于草本层和乔木层群落之间的相似性, 这说明冲沟沟头稳定过程中灌木层植物有趋同演化的趋势, 共有程度很高, 冲沟沟头活跃程度对灌木层植物群落功能造成的影响小于对草本层和乔木层植物群落。
| 表4 冲沟不同活跃程度植被群落相似(左下角)相异(右上角)系数矩阵 Table 4 The similarity (down left corner) and dissimilarity (upper right corner) index of the different gully |
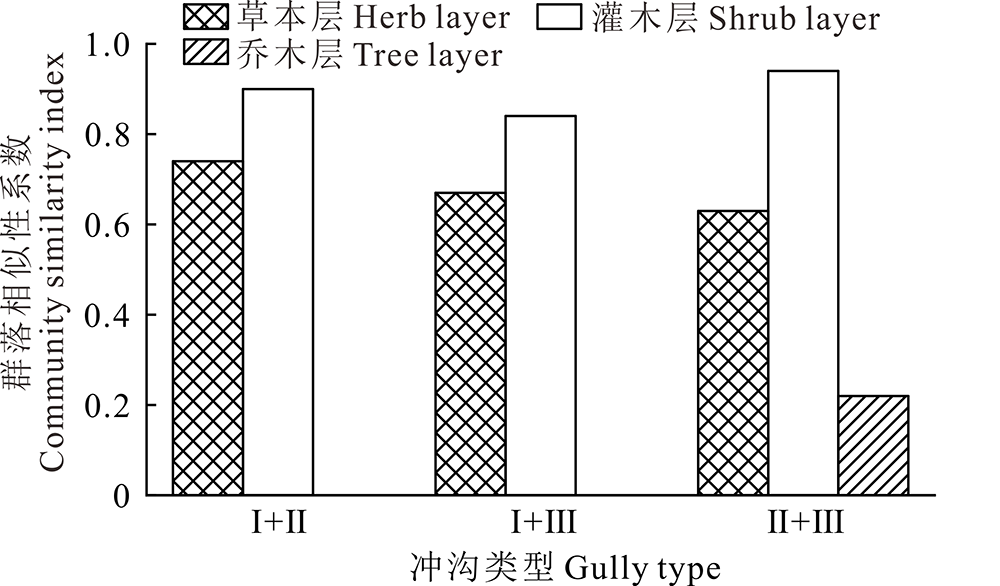 | 图2 不同活跃程度冲沟各层植被群落相似性系数Fig.2 The similarity coefficient of plant communities in different levels of gully at different degree of activity |
金沙江干热河谷区内冲沟分布广泛、沟头活跃程度不尽相同, 加之土壤肥力低下、砂砾含量高等诸多的因素, 制约着植物的系统发育过程, 形成了以稀树灌草丛为主的顶级植物群落结构类型[19]。本研究也发现, 不同活跃程度的冲沟沟头均以草本层的盖度最高, 其次是灌木层, 乔木层发育欠佳。
金沙江干热河谷冲沟发育区多年生草本植物占据草本层主要地位, 显著影响群落外貌和种类组成。而草本植物又以扭黄茅、孔颖草等为代表的禾本科植物和以柱花草、蔓草虫豆为代表的蝶形花科植物为主, 除柱花草外, 扭黄茅、孔颖草、蔓草虫豆等草本植物均为乡土物种, 在不同活跃程度的冲沟沟头样地中均有出现, 且在活跃型冲沟中仍具有覆盖面积大、长势良好的特点。这与王晨等[23]、杜敏晴等[24]的研究结果相同。主要原因在于干热河谷的水热条件极大程度地限制了外来物种, 而乡土植被在长期地演化过程中早已适应了高温缺水的恶劣环境; 且禾本科植物根系发达, 茎基部容易生出分蘖条, 而蔓草虫豆等蝶形花科植物能快速生长, 适应性强、易存活。因此, 在本区后续的植被恢复过程中应多考虑多年生禾本科和蝶形花科的草本植物, 可通过人工模拟飞播、人工撒播、人工封禁等措施促进植被恢复[16], 并且应充分发挥乡土植物适应性较强这一特点, 同时注重生物多样性, 避免发展成单一种优势群落, 导致植被群落稳定性降低, 抗干扰能力下降。
研究发现, 冲沟沟头的活跃程度对植物群落的数量特征具有显著影响。随着冲沟沟头逐渐趋于稳定, 草本层、灌木层植被种类、盖度逐渐增多, 具有较固定的植被组成, 植被多样性水平较高, 形成以草本搭配灌木为主的植物群落结构。这与杨振寅等[13]在元谋县所做的植物多样性的研究结果一致。其次, 冲沟沟头的活跃程度对灌木层和乔木层的生长情况的影响最为显著, 主要表现为:金沙江干热河谷冲沟发育区的乔木层发育欠佳, 但多样性及盖度随着冲沟沟头活跃度降低呈明显的增长趋势。这主要是植被对条件的反馈, 活跃型冲沟生境条件恶劣、水土流失严重, 乔木难以适应, 即使在一段时间内存活了, 也会在时间维度下逐渐淘汰。随着冲沟活跃度的降低, 以扭黄茅、孔颖草、车桑子、银合欢等为代表的适宜种能够快速生长, 可有效固土、增加土壤微生物含量和土壤养分含量, 改善局部土壤的理化性质, 营造较稳定的立地条件。其次植被盖度的提高和枯落物的增加可提高水源涵养能力、增加土壤入渗, 又可起到遮光隔热的作用以减少土壤水分的蒸发, 为乔木层的生长创造了相对较好的生境条件。
研究表明, 冲沟沟头的活跃程度可显著影响植物群落的多样性和群落结构的完整性及稳定性。随着冲沟沟头逐渐趋于稳定, 且其环境容量尚未达饱和的条件下, 植物多样性越来越高、层次性越来越明显、群落结构更加复杂和完善。对比不同活跃程度的冲沟, 稳定型冲沟的植被组成较半稳定型冲沟和活跃型冲沟更加完整。根据群落稳定性中的冗余理论[25], 稳定型冲沟植被备用种种类和个体数量较活跃型冲沟及半稳定型冲沟多, 群落的整体抗干扰能力要远强于活跃型冲沟的植物群落, 因此, 冲沟沟头的治理过程中应注意植被群落结构的完整性。
研究显示, 冲沟沟头的活跃程度对植被的优势种、先锋种无显著影响。因此, 在干热河谷冲沟发育区, 不同活跃程度的冲沟沟头, 植被恢复最优的植被配置均应是多年生禾本科草本植物搭配灌木。草本层以扭黄茅、孔颖草、三芒草最优, 灌木层以车桑子、银合欢最优。活跃冲沟植被覆盖度低, 且均以草本层为主, 但鉴于其恶劣的水肥条件难以为乔木的生长提供必需养分, 因而可不考虑乔木层, 主要利用鱼鳞坑、竹节沟、水平阶等造林技术进行灌木种植, 并撒播草种[16]。半稳定型冲沟植被覆盖度逐步升高, 草本层和灌木层群落结构较为完善、多样性指数较高, 主要需补种适宜乔木以提升植物群落功能及其稳定性。稳定型冲沟植被群落结构较复杂, 生态功能较为稳定, 不需人工配置植被, 依靠封禁等自然恢复措施即可。
1)不同活跃程度的冲沟不同层次植被群落优势种依次为扭黄茅、车桑子、印楝; 次优种为三芒草、孔颖草、银合欢、桉树, 其植被盖度随着冲沟稳定性的增加而增加, 表现为稳定型冲沟> 半稳定型冲沟> 活跃型冲沟。可将以上植被种类作为冲沟治理中的先锋物种进行种植。
2)金沙江干热河谷冲沟发育区植被种类随着冲沟稳定性的增加而愈加丰富。该区植物共有48种, 其中活跃冲沟内植物28种; 半稳定型冲沟内植物29种; 稳定型冲沟37种。不同冲沟草本植物均以禾本科、蝶形花科为主, 灌木以车桑子为主, 乔木层发育欠佳, 活跃型冲沟无乔木。
3)不同活跃程度的冲沟, 植被多样性高, 植被群落均以多年生草本搭配灌木为主, 乔木多样性水平较低, 活跃型冲沟甚至无乔木, 但随着冲沟稳定性的增加, 群落中原本处于劣势地位的乔木层多样性逐渐提升, 群落结构逐渐完善。
4)冲沟间稳定性差异越大, 植物群落相似性越低。而各层植被的群落相似性差异显著, 灌木层植物有趋同演化的趋势, 共有程度很高, 冲沟活跃程度对灌木层植物群落结构造成的影响小于对草本层和乔木层植物群落。活跃型冲沟、半稳定型冲沟、稳定型冲沟之间的植被群落相似程度表现为灌木层(极为相似)> 草本层(中等相似)> 乔木层(极度不相似)。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|