



{kind=link}
{kind=link}
{kind=link}
{kind=link}
短花针茅荒漠草原植被群落结构及生物量对水氮变化的响应
[李静 , 红梅
, 红梅* , 闫瑾, 张宇晨, 梁志伟, 叶贺, 高海燕, 赵巴音那木拉]
, 红梅, 闫瑾, 张宇晨, 梁志伟, 叶贺, 高海燕, 赵巴音那木拉]
|
|


作者简介:李静(1995-),女,内蒙古清水河人,在读硕士。E-mail: 1437325264@qq.com
为了解降水变化和氮沉降对荒漠草原植被群落结构和生物量的影响,在内蒙古四子王旗短花针茅荒漠草原进行水氮交互试验,采用裂区试验设计,主区为CK(自然降水)、W(增水30%)、R(减水30%),副区为0 (N0)、30 (N30)、50 (N50)、100 (N100) kg N·hm-2·a-1共12个处理。结果表明:1)水氮的介入导致植物群落组成、重要值发生了明显变化,R中群落物种数最低,提高了优势种在群落中的重要值,氮素添加导致群落物种数呈“抛物线”型,群落物种数先增多后减少。2) CK、W、R条件下随施氮量增加,物种丰富度、Shannon-wiener多样性指数和Pielou均匀度指数均先增大后减小,2016和2018年在CK-N30、W-N30、R-N50达到最大值,干旱年份(2017)分别在CK-N30、W-N50、R-N100达到最大值。水分主效应、氮素和水氮交互作用均对以上3个植物多样性指数有显著影响( P<0.05)。3)水分和氮素刺激了地上生物量的增加,短期氮素添加使多年生禾草生物量在地上总生物量中占比增加,多年生杂草生物量在地上总生物量中占比减小。水分、氮素、水氮交互均对地上生物量有显著影响( P<0.05)。
An experiment comprising a factorial combination of three water and four nitrogen levels (12 treatments), laid out in a split plot design, was conducted to understand the effects of precipitation change and nitrogen deposition on vegetation community structure and biomass in Stipa breviflora desert steppe in Siziwang Banner, Inner Mongolia. The main plot treatments were: CK (natural rainfall), W (30% rainfall addition), R (30% rainfall reduction), while the subplot treatments were 0 (N0), 30 (N30), 50 (N50), and 100 (N100) kg N·ha-1·a-1. The results show: 1) The application of water and nitrogen resulted in significant changes in community composition and species importance values. The R treatment had the lowest number of community species, and the importance value of dominant species in community was increased. The addition of nitrogen resulted in a curve of ‘parabola’ shape for number of species present (i.e. the number of community species was higher at medium nitrogen application rates than at N0 or N100). 2) With increased rate of nitrogen addition, species richness, Shannon-wiener diversity and Pielou evenness index were initially increased, but decreased at higher nitrogen application rates. The maximum values were observed in treatments CK-N30, W-N30 and R-N50 in 2016 and 2018, and the lowest value of CK-N30, W-N30, R-N50 in the drought year (2017), respectively. The main effects of water, nitrogen and water-nitrogen interaction all have significant effects on the above three plant diversity indexes ( P<0.05). 3) Water and nitrogen both stimulated aboveground biomass increase. Water, nitrogen as well as water and nitrogen interaction have significant effects on aboveground biomass ( P<0.05). The proportion of perennial grasses in the total aboveground biomass was increased by short-term nitrogen addition, while the percentage of perennial weeds was decreased.
降水和氮沉降作为全球变化的两个重要现象, 正不断地影响着陆地生态系统[1]。随着全球变化的加剧, 大气氮沉降迅猛增加, 预计在2050年达到195 Tg· a-1, 远远超出全球氮素临界负荷(100 Tg· a-1)[2, 3, 4]。高氮沉降会改变碳、氮、磷等元素的有效性, 进而影响植被生产力, 通过改变植物生态系统的组成来降低生物多样性[5, 6, 7]。同时, 降水量在全球尺度上也发生了明显变化[8], 根据联合国政府间气候变化专门委员会第五次气候变化评估报告显示, 1901年以来北半球中纬度陆地范围平均降水呈增加趋势[9]。降水是干旱、半干旱生态系统重要的水分来源, 降水格局变化从不同尺度影响植物生理及生态学过程[10, 11, 12], 尤其是受水分限制的荒漠草原生态系统, 由于物种数量相对较少, 外界细微的变化都很可能对生态系统造成很大的影响[13]。
在荒漠草原生态系统中, 氮沉降和降水变化带来的影响直接表现在植被群落结构中。目前, 国内外学者就单一氮沉降及水氮耦合对物种多样性的影响做了大量研究, 但得到的结论并不一致[14]。Zeng等[15]和Bai等[16]的研究都认为氮素添加后植物物种多样性降低, Huberty等[17]认为氮素添加对植物物种多样性无显著影响, 还有少数研究认为氮素添加使植物物种多样性增加[18]。多数研究都集中一次性施氮、设置单一施氮梯度或降水与集中一次性施入的氮素交互, 并且由于生态系统的多样性和地区之间的降水量差异, 试验结果往往不能直接应用到另一个生态系统中[19]。
为了更好地解释氮沉降和降水对荒漠草原植被结构及多样性影响的变化, 本研究以内蒙古乌兰察布市四子王旗短花针茅(Stipa breviflora)荒漠草原为研究对象, 探究水分、氮素及水氮交互对荒漠草原群落结构、物种多样性和生物量的影响。现提出以下科学问题:1)短花针茅荒漠草原降水量低, 养分贫瘠, 水分、氮素、水氮交互将如何影响荒漠草原群落结构及多样性; 2)水分、氮素、水氮交互对荒漠草原植被生物量有何影响, 是否会限制植物的生长。研究大气氮沉降和降水格局变化对荒漠草原生态系统的影响, 以期对荒漠草原生态系统提供科学的管理对策, 实现其可持续发展。
试验区位于内蒙古乌兰察布市四子王旗短花针茅荒漠草原生态系统野外科学试验基地, 地理坐标为111° 53' E, 41° 47' N, 海拔1450 m, 属于温带干旱、半干旱大陆性季风气候区, 年平均降水量为280 mm, 主要集中在6-9月, 年平均气温为3.4 ℃。2017年全年降水同往年相比较少, 降水量为258.8 mm, 降水主要集中在6、7、10月, 气温同往年相比有所上升, 平均气温为5.65 ℃(图1)。该试验区地带性植被是短花针茅荒漠草原, 植物种类较少, 主要优势种有短花针茅和无芒隐子草(Cleistogenes songorica)。
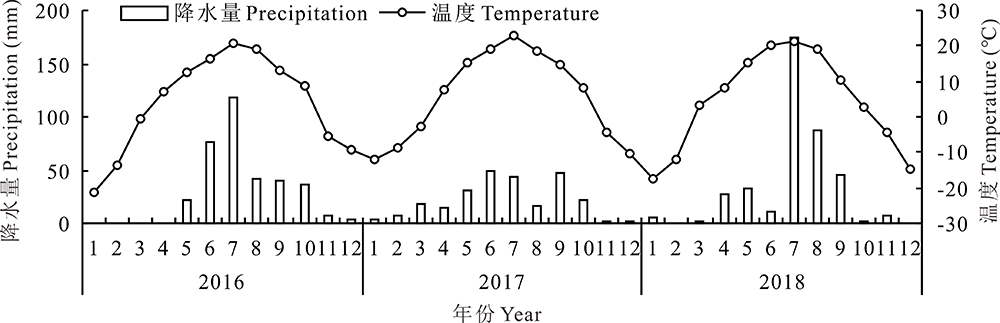 | 图1 3年气温和降水动态Fig.1 The temperature and precipitation patterns in three years |
试验于2015年12月在样地围栏内进行, 采用裂区设计, 主区为3个水分处理, 分别是自然降水(CK)、增水30%(W)和减水30%(R), 副区为4个氮素处理, 分别是0 (N0)、30 (N30)、50 (N50)、100 (N100) kg N· hm-2· a-1(不包括大气沉降的氮量)共12个处理, 即:对照(CK-N0)、自然降水施氮30 kg N· hm-2· a-1(CK-N30)、自然降水施氮50 kg N· hm-2· a-1(CK-N50)、自然降水施氮100 kg N· hm-2· a-1(CK-N100)、增水不施氮(W-N0)、增水施氮30 kg N· hm-2· a-1(W-N30)、增水施氮50 kg N· hm-2· a-1(W-N50)、增水施氮100 kg N· hm-2· a-1(W-N100)、减水不施氮(R-N0)、减水施氮30 kg N· hm-2· a-1(R-N30)、减水施氮50 kg N· hm-2· a-1(R-N50)、减水施氮100 kg N· hm-2· a-1(R-N100)。增水试验分别在每年5-8月的1-3日进行, 通过对荒漠草原多年的降水量监测并做模型预测未来降水量增减极限, 增加量为近5年5月(18.4%)、6月(17.0%)、7月(28.3%)、8月(36.3%)平均降水量的30%, 减水试验通过减水架减少当地年平均降水量的30%, 减水架由条状凹槽式亚克力板构成, 亚克力板遮挡面积占小区面积的30%达到减水30%。根据本地区氮沉降情况, 氮处理强度和频度参考国际上同类研究的处理方法, 施氮处理按照施氮量换算成小区硝酸铵(NH4NO3)施用量, 为了尽可能均匀施氮, 在每年生长季(5-9月)每月一次, 将每个小区每次施用硝酸铵的量溶于30 L水中(在增水处理之后), 均匀喷洒在每个小区内, 非生长季(10月-翌年4月), 将每月每个小区施氮量与风干土(直径< 2 mm)按肥土比1∶ 10充分混匀, 在无风时以模拟氮沉降的方式直接撒施。试验共12个处理3次重复, 共计36个样方, 每个小区面积为7 m× 7 m=49 m2, 各小区间设置2 m隔离带。
野外调查工作于每年植物生长旺季8月中旬进行, 在各小区内随机选择1 m× 1 m样方, 再将样方分成100个10 cm× 10 cm的小方格, 测量并记录样方内各植物种的高度、密度、盖度、频度。生物量的测定为在各小区内随机设置0.5 m× 0.5 m的小样方, 之后采用收割法齐地剪取植物地上部分装入信封中, 每个小区重复3次, 带回实验室在65 ℃恒温箱烘干24 h称重。将样方内物种根据生活型分成4个类群:一、二年生植物、多年生杂草、多年生禾草和半灌木、小半灌木。
物种的相对重要值=(相对密度+相对高度+相对干重)/3[20]
地上生物量用植物干重表示。物种丰富度(P)用单位面积出现的物种数(S)表示:P=S。
Shannon-wiener指数[21]:
Pielou均匀度指数[22]: E=H'/lnS
式中:Pi为第i个物种的个体数占总个体数的比率; S为物种数。
采用SAS 9.1软件对地上生物量、植物多样性指数进行单因素方差分析(One-way ANOVA)并对植物多样性指数做双因素方差分析(Two-way ANOVA), 用邓肯多重比较检验(Duncan’ s multiple range test)在方差分析中有差异的变量间的差异显著。采用Microsoft Excel 2003进行制图。
2016年植被群落有刺藜(Chenopodium aristatum)、猪毛菜(Salsola collina)、灰绿藜(Chenopodium glaucum)、猪毛蒿(Artemisia scoparia)、 栉叶蒿(Neopallasia pectinata)、 银灰旋花(Convolvulus ammannii)、 阿尔泰狗娃花(Heteropappus altaicus)、苔草(Carex tristachya)、短花针茅、无芒隐子草、克氏针茅(Stipa krylovii)、蒙古韭(Allium mongolicum)、细叶韭(Allium tenuissimum)、冷蒿(Artemisia frigida)、木地肤(Kochia prostrata), 与CK-N0相比, 增水施氮(W-N)、CK中N30、N50、N100和减水施氮(R-N)R中N50、N100的物种数目有增加趋势, 而R-N0和R-N30的物种数目减少。2017年荒漠草原的群落组成依旧是由一、二年生植物、多年生杂草、多年生禾草和半灌木、小半灌木组成, 但物种数减少。2018年同2016年相比, 一、二年生植物在群落中的比例增加, 多年生杂草减少, 其中阿尔泰狗娃花和苔草消失, 刺藜、灰绿藜、猪毛蒿和银灰旋花在群落中占比增加。
水、氮作用下, 物种的重要值不同, 其中多年生禾草短花针茅的重要值最高, 其次是无芒隐子草。2016年, 在W中随着氮素的添加, 猪毛菜的重要值逐渐减小, 短花针茅先减小后增大, 木地肤逐渐增大。在CK中随氮素的添加, 猪毛菜的变化规律不明显, 栉叶蒿在N0的重要值最大, 短花针茅重要值先减小后增大。R提高了短花针茅、克氏针茅和细叶韭的重要值。2017年小半灌木木地肤发育良好, 重要值随氮素添加逐渐增大。随着时间的延长, 2018年群落中刺藜、灰绿藜、猪毛蒿和银灰旋花所占比例增加, 在W中随氮素添加, 刺藜重要值先减小后增大, 灰绿藜和猪毛蒿分别在N100、N0达到最大值, W处理提高了栉叶蒿的重要值。在CK中, 刺藜重要值随氮素添加先减小后增大, 在N0、N100达到最大值。在R中, 刺藜重要值随氮素添加先增大后减小, 在N50达到最大值(表1)。
| 表1 不同年份水氮添加对短花针茅荒漠草原植物种群重要值的影响 Table 1 Effects of water and nitrogen addition on important value of different population in S. breviflora desert steppe in different years |
2016年在3种不同水分条件下随着氮素的添加, 物种丰富度、Shannon-wiener指数、Pielou均匀度指数的变化趋势相同, 均呈先增大后减小, 分别在W-N30、CK-N30、R-N50达到最大值, 但均无显著差异。各指数在W-N30处理下分别增加了16%、8%、6%, CK-N30处理下分别增加了10%、5%、6%, R-N50处理下物种丰富度、Shannon-wiener指数分别增加了7%、6%, 均匀度指数无变化。在相同氮素添加条件下, 各水分处理中物种丰富度、Shannon-wiener指数均为W> CK> R, 均匀度指数变化幅度较小。2017年在3种不同水分条件下, 随着氮素的添加, 物种丰富度、Shannon-wiener指数、Pielou均匀度指数呈先增大后减小的趋势, 分别在W-N50、CK-N30达到最大值, 在R处理中随着氮素的添加, 各指数呈逐渐增大的趋势, 均在N100达到最大值, 且对各指数均无显著影响。在相同氮素添加下, 物种丰富度、Shannon-wiener指数、Pielou均匀度指数均呈W> CK> R且W对群落多样性有显著影响(P< 0.05)。2018年各指数变化趋势和2016年相同, 在3种不同水分条件下, 随着氮素的添加物种丰富度、Shannon-wiener指数、Pielou均匀度指数同样呈先增大后减小的趋势, 分别在W-N30、CK-N30和R-N50达到最大值, 各指数在W-N30处理下分别增加了21%、17%、16%, CK-N30处理下分别增加了6%、13%、16%, R-N50处理下分别增加了9%、15%、15%。相同氮素添加不同水分处理中, 物种丰富度、Shannon-wiener指数和均匀度指数为W> CK> R, 增水对物种丰富度和Shannon-wiener指数有显著影响(P< 0.05)。2016年同2018年相比, 各处理下物种丰富度和Shannon-wiener指数变化较小, 但均匀度指数有减小的趋势(图2)。
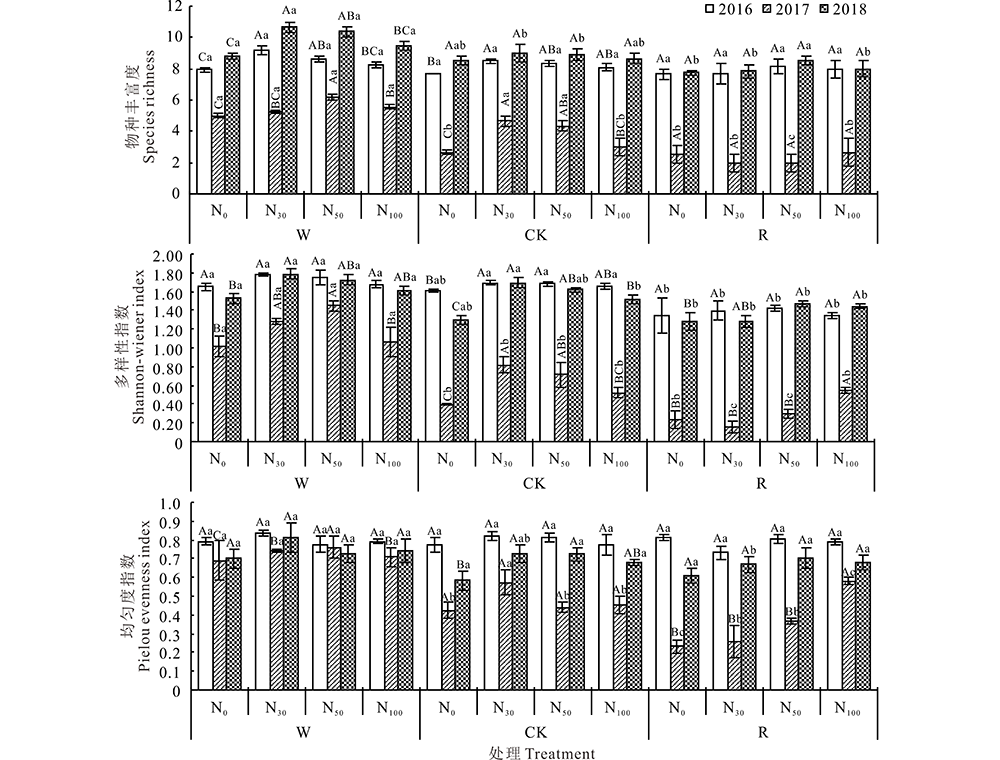 | 图2 不同年份水氮添加对物种丰富度、Shannon-wiener指数和Pielou均匀度指数的影响 W: 增水30%; CK:自然降水; R:减水30%。不同大写字母表示同一水分处理下不同氮素水平间差异显著(P< 0.05), 不同小写字母表示同一氮素处理下不同水分处理间差异显著(P< 0.05), 下同。Fig.2 Effects of water and nitrogen additions on species richness, Shannon-wiener index and Pielou evenness index in different years W: 30% rainfall addition; CK: Natural rainfall; R: 30% rainfall reduction. Different capital letters showed significant difference among different nitrogen levels under the same water treatment (P< 0.05), different small letters showed significant difference among different water treatments under the same nitrogen treatment (P< 0.05), the same below. |
为了明确水、氮及水氮交互作用对植被群落多样性的影响, 进行了双因素方差分析(表2)。综合分析各处理间的物种丰富度、Shannon-wiener指数、Pielou均匀度指数表现为2017年最低, 而2016和2018年大致相同。水氮双因素方差分析结果表明, 水分、氮素及水氮交互作用均对物种丰富度和植物群落多样性有显著影响, 相比氮素, 水分的作用更大。
| 表2 水氮添加对荒漠草原物种多样性影响的双因素方差分析 Table 2 Two way analysis of variance for the effects of water and nitrogen addition on species diversity in desert steppe communities |
3年综合分析, W、CK、R条件下, 随着氮素的添加地上生物量呈逐渐增大的趋势, 分别在W-N100、CK-N100、R-N100达到最大值。CK中N30、N50、N100分别增加了14.35%、20.05%、24.15%, W中N30、N50、N100分别增加了16.69%、21.49%、31.72%, R中N30、N50、N100分别增加了8.00%、10.50%、15.90%。相同氮素添加下, 地上生物量在3种不同的水分处理下表现为W> CK> R, 增水对地上生物量有显著影响(P< 0.05), 和CK-N0相比较, W-N0增加了17.26%, R-N0减少了32.46%(图3)。
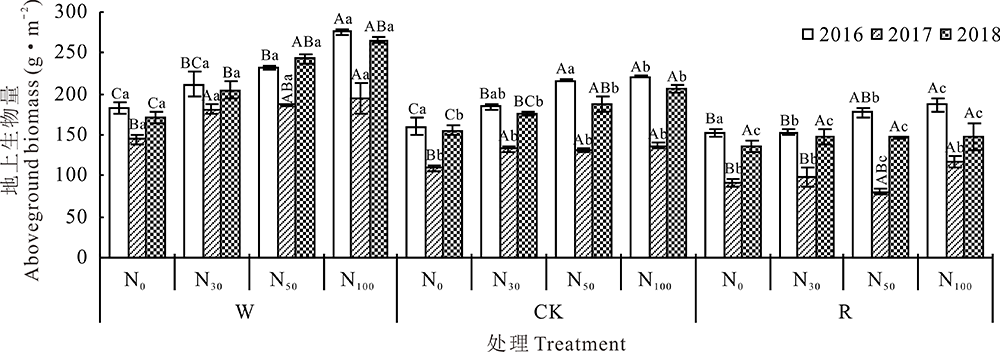 | 图3 不同年份水氮添加对群落地上总生物量的影响Fig.3 Effects of water and nitrogen additions on total aboveground biomass in different years |
地上生物量按植物功能型表现为多年生禾草> 一、二年生植物> 多年生杂草> 半灌木、小半灌木。一、二年生植物和多年生禾草在各水分处理中随着氮添加, 地上生物量显著增加(P< 0.05), 且在总地上生物量中的占比均随着氮素添加量的增多而逐渐增大。多年生杂草在W中随氮添加生物量有增加的趋势, CK、R中随着氮素添加生物量有减小的趋势, 其在总地上生物量中的占比随着氮素添加量的增多而逐渐减小。相同氮素添加条件下, 各功能型植物生物量均呈W> CK> R。2017属干旱年份, 各功能群植物生物量均较低。群落地上总生物量在年际间(2016、2018年)无明显差异, 但多年生杂草、多年生禾草生物量在群落地上总生物量中占比有所变化, 多年生杂草生物量占群落地上总生物量的比例从9.78%下降到3.58%, 多年生禾草生物量在群落地上总生物量中的占比从63.75%增加到68.30%, 但在2017年, 半灌木、小半灌木的生物量相较于2016和2018年最高(图4)。为了明确水氮控制对生物量的影响, 进行了水氮双因素方差分析。结果表明, 水分、氮素以及水氮互作均对短花针茅荒漠草原生物量有显著影响(P< 0.05), 其中水分是主要限制因子(表3)。
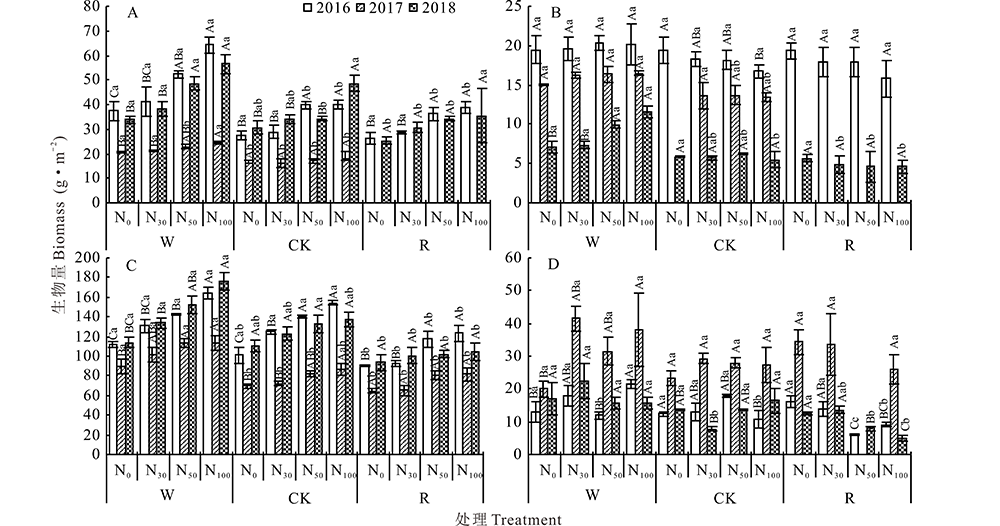 | 图4 不同年份水氮添加对各功能型植物生物量的影响 A:一、二年生植物Annual and biennial plants; B:多年生杂草Perennial weeds; C:多年生禾草Perennial grass; D:半灌木、小半灌木Semi-shrub and undershrub.Fig.4 Effect of water and nitrogen addition on biomass of functional plants in different years |
| 表3 水氮添加对荒漠草原生物量影响的双因素方差分析 Table 3 Two way analysis of variance for the effects of water and nitrogen addition on biomass in desert grassland communities |
短花针茅荒漠草原植物群落由一、二年生植物、多年生杂草、多年生禾草和半灌木、小半灌木组成。年际间群落组成变化大, Yang等[23]发现降水增加和氮添加显著改变了功能组水平的植物群落结构和组成, 植物群落结构是在不断变化的环境条件下由水介导的。张彦东等[24]认为在退化严重的草地上施加氮肥, 禾本科草本植物大量增加, 乔木灌木密度增加不明显, 本研究与其具有相似规律。本研究结果表明, 多年生禾草植物在群落中始终占绝对优势, 试验的第3年猪毛菜减少, 藜科类植物(刺藜、灰绿藜)在群落组成中所占比例增大, 半灌木、小半灌木对水氮控制变化的影响不明显, 藜科大多数种类是盐生、沙生和喜氮植物[25], 水分的增加和连续施氮的积累使藜科在群落中迅速生长。相同氮素添加, 增水条件下植物群落组成数量大于其他两个水分处理, 使优势物种的重要值降低, 这与陶冶等[26]在新疆北部荒漠的研究结果一致, 是由于向环境中输入了水分使群落物种丰富度增加, 促进了各植物种的生长, 尤其是对水分敏感的一、二年生植物。增水条件下, 短花针茅对氮素添加的反应最明显, 随着氮素的增加短花针茅在群落中的重要值呈先减小后增大的趋势, 而在自然降水和减水处理中重要值变化不明显, 适量的氮素添加可以满足植物生长的需求, 荒漠草原干旱少雨, 群落特征对水分条件敏感, 在有水分的前提下植物能更好地吸收氮素[27], 使得其他植物在群落中增多, 导致短花针茅重要值反而先减小, 因此, 水分和氮素添加对群落组成及重要值产生了明显影响。Tilman[28]的生态位优先占领假说指在生境较差且物种数相对较少的群落, 第一位优势种首先占领生态位空间的大部, 第二位的占领其余下的大部空间, 依此类推, 到末位的只能占留下的很少空间, 本试验虽更趋向于Tilman生态位优先占领假说, 但由于天然草原植物群落的复杂性, 水分变化、氮素添加以及水氮互作对群落结构及组成的影响机理还需进一步探究。
本试验中, 在相同氮素添加条件下, 各指数均呈W> CK> R, 说明水分添加利于群落多样性增加, 与白春利等[27]的研究结果相似。在3种水分条件下, 施入氮肥的第一年, 物种丰富度、Shannon-wiener多样性指数和Pielou均匀度指数均随着氮素的添加表现为先增加后下降的趋势, 但无显著差异(P> 0.05)。因生态系统对环境变化有缓冲作用, 所以短期氮素添加并没有引起群落多样性的明显变化, 这与赵新风等[29]在施肥和增水第一年的结果相似。施入氮肥的第二年, 各指数随氮素添加先上升后下降, 氮素添加对物种丰富度、Shannon-wiener多样性指数有显著影响(P< 0.05), 对Pielou均匀度指数的影响不显著(P> 0.05)。各指数在2018年也表现为随氮素添加先上升后下降, 分别在CK-N30、W-N30、R-N50达到最大值, 之后显著下降。Stevens等[30]也认为氮素的增加会使物种丰富度降低, 是由于嗜氮植物对氮的需求增加导致的。Theodose等[31]认为在贫瘠环境下, 物种多样性有增值空间, 短时间内不发生竞争淘汰, 而在肥沃的环境下, 养分添加后物种多样性增值的空间不大, 物种间产生竞争导致一些物种被淘汰。本研究分析出现此结果的原因:1)氮素的添加使群落多样性先升高后降低, 在低氮添加时, 土壤养分增多, 植物可利用氮素随之增大, 随着氮素的增加, 各物种表现出种间竞争, 一部分植物对氮素吸收能力强, 造成其他植物竞争能力下降, 最终导致单一植物生长迅速, 群落多样性下降; 2)氮素添加增加了土壤中的有效养分, 促进了荒漠草原地上部分优势种群的生长[32], 地上生物量的增加导致竞争排斥[33], 例如多年生禾草(短花针茅、无芒隐子草)增加, 植株较高的禾草类植物对于光的竞争排斥使植株矮小的植物被淘汰[34]; 3)群落多样性指数的变化除与植被本身特性有关外, 还可能受非生物条件影响, 例如土壤酸碱度和其他资源可用性会降低物种丰富度和多样性指数[25]。
本研究中, 各水分条件下随着氮素的添加群落地上总生物量增大, 氮素添加为荒漠草原植物的生长提供了养分需求, 增加了群落地上生物量。在相同氮素添加条件下, 增水处理为土壤提供更多的水分, 为植物生长提供充足的水源, 促进植物营养生长, 使得地上生物量得到提高, 这与郭蕊等[35]的研究结果一致。郭永盛[36]认为水氮添加使土壤中速效养分增加, 植物功能群得到有效的可利用资源。半灌木、小半灌木生物量随着水分变化不会大幅波动, 一、二年生植物和多年生草本植物生物量受水分影响较大, 这与孙岩等[37]的研究结果一致, 深根系的灌木主要利用深层土壤水分, 因此, 生物量不会发生大幅波动, 而浅根系一年生或多年生草本主要利用浅表层土壤水分, 其生物量的积累和浅表层土壤水分密切相关。除此之外, 功能群间的相互制约和功能群自身特性也会受到影响[38]。各植物功能群中, 多年生禾草占群落地上总生物量比例在第3年增加, 而多年生杂草占群落地上总生物量比例减小, 多年生禾草对水氮添加的响应较显著, 因其大多植株高大, 地上部分生长迅速, 高度和盖度增加, 处于光养分竞争的最上层, 有资源竞争优势, 且其根系是须根系, 对水分和养分添加反应敏感[39], 而多年生杂草主要由菊科类植物组成, 随着氮素增加因种群植物竞争加大而使其地上生物量减少, 氮素的添加缓解了其对多年生禾草植物的限制, 植物竞争格局因养分限制的消除发生变化, 进而群落结构发生改变[38]。
1)在水氮控制的影响下, 植物群落组成、重要值发生了明显变化, R中群落物种数最低, 提高了优势种在群落中的重要值, 施氮量增加导致群落物种数呈“ 抛物线” 型, 群落物种数先增多后减少, 猪毛蒿、银灰旋花、木地肤以及多年生禾草具有较强的竞争力, 在群落组成中占优势地位。
2)CK、W、R条件下随施氮量增加, 物种丰富度、Shannon-wiener多样性指数和Pielou均匀度指数均先增大后减小, 2016和2018年在CK-N30、W-N30、R-N50达到最大值, 干旱年份(2017年)分别在CK-N30、W-N50、R-N100达到最大值。水分主效应、氮素和水氮交互作用均对以上3个植物多样性指数有显著影响(P< 0.05)。
3)水分和氮素刺激了地上生物量的增加, 短期氮素添加使多年生禾草生物量在地上总生物量中占比增加, 多年生杂草生物量在地上总生物量中占比减小, 半灌木、小半灌木受水氮控制的影响较小。水分、氮素、水氮交互均对地上生物量有显著影响(P< 0.05)。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|