


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
围封和防除狼毒对狼毒斑块土壤理化性质和微生物量影响的研究
[鲍根生*  , 宋梅玲, 王玉琴, 尹亚丽, 王宏生]
, 宋梅玲, 王玉琴, 尹亚丽, 王宏生]
, 宋梅玲, 王玉琴, 尹亚丽, 王宏生]
|
|


作者简介:鲍根生(1980-),男,青海乐都人,助理研究员,博士。E-mail: baogensheng2008@hotmail.com
狼毒是青藏高原高寒退化草地主要滋生的毒害草种类,采用化学除草剂和草地围封方法能有效遏制狼毒种群扩张的现状。然而,针对狼毒形成微斑块对使用除草剂和草地围封措施后植被群落结构、土壤理化性质和土壤微生物量响应变化的研究较少。基于此,狼毒型退化草地采用短期围栏封育(1年),以围封地未防除和除草剂处理的狼毒微斑块为研究对象,比较和分析狼毒斑块内外物种多样性指数、土壤养分和土壤微生物量变化。结果表明:1)围封地未防除狼毒斑块内生物量和多样性指数高于狼毒斑块外,而防除狼毒斑块外生物量高于狼毒斑块内。2)围封地未防除狼毒斑块内表层土壤的全氮、硝态氮、铵态氮、速效磷、有机质和土壤含水量高于狼毒斑块外,而防除狼毒斑块外表层土壤的有机质和全钾含量高于狼毒斑块内;围封地未防除狼毒斑块内深层土壤铵态氮和硝态氮含量高于斑块外,防除狼毒斑块外深层土壤速效磷、铵态氮和有机质含量高于斑块内。3)防除狼毒和围封地未防除狼毒斑块外土壤微生物量碳高于狼毒斑块内,防除狼毒斑块外土壤微生物量氮增加而微生物量磷却快速降低。由此可见,狼毒型退化草地治理需采用“狼毒净”化学除草剂快速降低狼毒种群并辅以长期围封(禁牧)措施,可提高禾本科等优良牧草比例、调节草地群落结构趋于稳定和加速土壤养分循环,并最终实现狼毒型退化草地生态修复的目标。
Stellera chamaejasme is a toxic plant, largely distributed in degraded grassland of the Qinghai-Tibetan Plateau, where it can dominate other vegetation. Grazing exclosure and herbicides are considered the effective methods to control S. chamaejasme and restore degraded grassland where S. chamaejasme has become the dominant species. However, effects of grazing exclosure and herbicides on plant community structure, soil physical-chemical properties and soil microbial biomass within or between patches of S. chamaejasme are unclear. Therefore, plant diversity indexes, soil nutrient status, and soil microbial biomass were determined within and between S. chamaejasme patches after one year of grazing exclosure and herbicide application. It was found that aboveground biomass and plant diversity indexes within natural patches of S. chamaejasme were higher than between patches; however, aboveground biomass within herbicide application patches was lower than between them. In the surface soil (0-10 cm), Stellera increased soil moisture content, total nitrogen, ammonium, nitrate and organic matter. By contrast, herbicide decreased the organic matter and total potassium within patches. At greater soil depth (10-20 cm), Stellera increased ammonium and nitrate content; in contrast, herbicide decreased available phosphorus, ammonium and organic matter. Herbicide and grazing exclosure increased microbial carbon biomass within patches compared with between patches. Furthermore, herbicide increased soil microbial nitrogen biomass but decreased microbial phosphorus within patches. Combing herbicide and grazing exclosure decreased the S. chamaejasme population, improved grass biomass, regulated plant community structure and accelerated nutrient cycling, and resulted in restoration of the degraded grassland in which S. chamaejasme had become the dominant species.
狼毒(Stellera chamaejasme), 别名断肠草、馒头花和瑞香狼毒, 属瑞香科狼毒属多年生草本植物[1]。狼毒属超旱生植物, 具有粗长的轴状根系, 能适应极端干旱和寒冷生境, 成为高寒草原和荒漠草原植被的优势种[2, 3]。狼毒植株体内合成的多种次生代谢物, 尤其是黄酮类化合物, 对草食家畜、昆虫和病原菌具有毒害作用, 导致草食家畜对狼毒取食行为表现出明显的趋避性[4]。另外, 全球气候暖化、草地载畜量持续过高和啮齿动物破坏等因素的综合作用, 导致狼毒在青藏高原退化草地中大面积出现, 成为退化草地主要优势物种, 严重影响当地草地畜牧业的健康发展[5, 6]。
狼毒作为青藏高原退化草地危害最严重的毒害草种类之一, 以往研究主要集中于狼毒防除、狼毒次生代谢物提取和狼毒种群生态学等方面[7, 8, 9, 10]。目前, 狼毒防除主要采用化学防除和人工拔除等方法, 化学防除能短期内实现遏制狼毒大面积蔓延的现状, 但施用化学除草剂易对其他阔叶类植物造成药害, 进而降低草地植物多样性和群落稳定性[7]; 人工拔除狼毒能克服化学除草剂对草地植被和群落的不利影响, 但防治效率较低[11]。另外, 狼毒种群学研究通过调查狼毒分布区域和分析草地群落组成影响, 已明确狼毒种群扩张趋势和对草地植物群落结构的影响[12, 13]。然而, 针对狼毒植株对同域其他植物分布格局和土壤养分循环影响方面的研究报道较少[14, 15]。
草地围封成为青藏高原退化草地原生植被恢复的最佳治理措施之一[16], 通过禁牧能提高禾草等可食牧草比例和加速土壤矿物质循环周期[17, 18, 19]。Wu等[17]研究发现以藏嵩草(Kobresia tibetica)为优势种的高寒草甸, 经5年封育后禾本科和莎草科植物盖度显著提高, 而毒害草盖度显著降低, 同时土壤氮素和磷素含量显著增加。Shang等[20]研究也证明高寒草地封育3年后禾本科植物生物量显著增加, 而杂类草盖度快速降低。王玉琴等[21]研究报道狼毒型退化草地采用封育措施后禾本科和莎草科生物量增加58%, 而狼毒盖度降低60%。可见, 草地封育措施通过限制草食家畜对可食牧草采食频率, 为可食牧草提供休养生息周期, 进而提高可食牧草竞争能力并抑制毒害草种群扩张, 最终实现毒害草型退化草地生态修复目标。
草地围封联合使用化学除草剂技术可能成为现阶段针对狼毒型退化草地最有效的治理措施[21, 22, 23], 上述研究仅从喷施化学除草剂防除狼毒或使用草地围封等单项技术研究狼毒大面积发生对草地群落结构和土壤理化性质的影响, 然而将狼毒防除和草地围封技术耦合, 以单株狼毒形成微斑块为切入点, 研究狼毒防除和草地围封耦合技术对狼毒斑块内草地群落结构、土壤养分和土壤微生物量影响的研究鲜有报道。基于此, 本研究以单株狼毒形成的微斑块为研究对象, 在草地围封条件下采用化学除草剂防除对狼毒微斑块进行扰动, 并进行围封区防除和未防除狼毒斑块内外对比研究, 主要解释以下问题:1)围封条件下狼毒防除对狼毒斑块内外植物生物量和多样性指数的影响; 2)围封条件下狼毒防除对狼毒斑块内外土壤理化性质和土壤微生物量变化的影响。通过解答上述问题, 可明晰狼毒微斑块的生态功能和特征, 这将为狼毒型退化草地的综合治理提供基础数据。
试验地位于青海省海北藏族自治州海晏县青海湖乡达玉德吉村(N 37° 04'01″, E 100° 52'48″)。该区域气候类型为高原大陆性气候, 冬季漫长, 夏季短暂。降水集中于5-9月, 年降水量为388.6 mm。试验地为当地牧民夏季牧场, 每年6-10月中旬草食家畜在样地周围进行自由采食, 放牧强度为4只藏羊· hm-2。由于草地载畜量持续过高导致草地狼毒大面积发生, 密度高达8株· m-2。
试验地草地类型为高寒草甸草原, 草地建群种为线叶嵩草(Kobresia capillifolia)和羊茅(Festuca ovina), 优势种为狼毒, 伴生禾草为垂穗披碱草(Elymus nutnas)、草地早熟禾(Poa pratensis)和异针茅(Stipa aliena), 其他阔叶类植物为秦艽(Gentiana macrophylla)、山莓草(Sibbaldia adpressa)、西藏附地菜(Trigonotis tibetica)、扁蓿豆(Melilotodes ruthenica)和急弯棘豆(Oxytropis deflexa)等。试验区域土壤类型为高山草甸土。
2015年7月通过实地调查发现, 试验地为狼毒高密度发生区(平均密度5株· m-2), 狼毒株丛平均间距为30 cm, 单株狼毒冠幅为25~30 cm。2016年6月狼毒开花初期, 随机建立4块面积为6400 m2 (80 m× 80 m)的围封区。依据邢福等[24]的研究结果, 在围封区内狼毒分布均匀地段选取40单株狼毒为研究对象, 选择标准为单株狼毒枝条数20~25条(生长年限≥ 5年), 并在选取狼毒基部2~3 cm处用木桩标定位置。依据前期狼毒冠幅测定, 以狼毒基部为圆点, 直径30 cm处划定为狼毒斑块内, 将两株狼毒斑块内以外区域定义为狼毒斑块间。7月下旬, 狼毒盛花期挑选20株狼毒采用青海省畜牧兽医科学院研制的“ 狼毒净” 进行防除[7], “ 狼毒净” 有效浓度为1.125 L· hm-2; 剩余木桩标记的狼毒定义为围封区未防除狼毒。2017年6月, 牧草返青期对木桩标记点防除狼毒返青率调查发现, 狼毒返青率仅为1%。
1.2.1 狼毒斑块内(间)植物组成调查 2017年8月牧草开花期, 根据标记木桩在围封区随机选择10株防除和未防除狼毒, 以木桩为圆点, 直径30 cm样圆调查狼毒斑块内出现植物频度、盖度和生物量。同时, 按照上述狼毒斑块外划定方法, 采用狼毒斑块内植物调查方法对狼毒斑块外植物组成进行调查。通过下述植物重要值计算方法计算样圆内出现植物的重要值[17]。
重要值(important value, IV)=(相对频度+相对盖度+相对地上生物量)/3
依据样圆内出现植物重要值, 计算香农威纳指数(H)、优势度指数(D)和均匀度指数(E), 具体计算公式如下:
香农威纳指数:
物种优势度指数:
均匀度指数:
式中:S代表样圆内出现的植物种类数量, Pi=ni/N表示第i个物种的相对多度, ni代表第i个植物个体数量, N表示样圆内出现植物的个体总数。
1.2.2 狼毒斑块内外土壤理化性质和土壤微生物量分析 围封区选取防除和未防除狼毒斑块各10个, 用孔径4 cm土钻分别钻取斑块内外0~20 cm土壤样品。以10 cm深度为标准, 将土样分成0~10 cm和10~20 cm两层。其中, 按照相同试验设计和同一土层深度的标准, 随机将3个样点土壤进行混匀。其中, 土壤含水量、pH、有机质、全氮、全磷、全钾、速效磷、速效钾、铵态氮和硝态氮测定方法参考鲍士旦[25]《土壤农化分析》方法。在防除和未防除狼毒斑块内外用土钻钻取0~20 cm深度土壤, 5份土样混合成1份, 每个处理土样重复3次, 采用氯仿熏蒸提取-重铬酸钾容量法测定土壤微生物量碳含量[26], 半微量蒸馏法测定土壤微生物量氮含量[27], 钼锑抗比色法测定土壤微生物量磷含量[28]。
采用一般线性模型方法分析防除和未防除狼毒斑块内外地上生物量、多样性指数、土壤理化性质、土壤微生物量碳、土壤微生物量氮和土壤微生物量磷变化; 其中, 狼毒防除处理和斑块内(外)为固定因子, 不同狼毒株丛为区组。检验固定因子与区组间交互效应; 另外, 采用Tuckey检验分析防除和未防除狼毒斑块内外测试指标的差异性。利用SPSS 22.0软件进行数据分析, 图中数据采用平均值± 标准误表示。
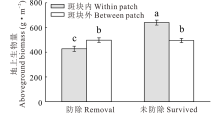
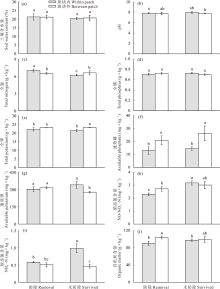
草地围封条件下, 防除狼毒处理斑块外生物量显著高于狼毒斑块内, 而未防除狼毒斑块内生物量显著高于狼毒斑块外(P< 0.05, 图1)。同时, 围封区未防除狼毒斑块多样性指数(包括香农威纳指数、优势度指数和丰富度指数)显著高于防除狼毒斑块(P< 0.05, 图2)。其中, 未防除狼毒斑块内多样性指数显著高于狼毒斑块外和防除狼毒斑块内外(P< 0.05, 图2); 防除狼毒斑块内外香农威纳指数和优势度指数差异不显著(P> 0.05, 图2a, b), 而防除狼毒斑块内丰富度指数显著高于防除狼毒斑块外(P< 0.05, 图2c)。
 | 图1 围封条件下未防除和防除狼毒斑块内外地上生物量变化 不同小写字母表示防除和不防除狼毒斑块内外差异显著(P< 0.05), 下同。Fig.1 Biomass of natural surviving and removing treatment within or between patch of S. chamaejasme in enclosed plot Different lowercases indicate the significant difference of natural surviving and removing treatment within or between patch of S. chamaejasme in enclosed plot, the same below. |
 | 图2 围封条件下未防除和防除狼毒斑块内外草地群落多样性指数变化Fig.2 Plant diversity indexes of natural surviving and removing treatment within or between patch of S. chamaejasme in enclosed plot |
草地围封条件下, 防除狼毒能显著影响狼毒斑块内外土壤理化性质(图3和图4)。其中, 未防除和防除狼毒斑块内表层土壤(0~10 cm)含水量显著高于狼毒斑块外, 防除狼毒斑块内土壤含水量显著高于未防除狼毒斑块内和斑块外(图3a); 而狼毒斑块内外深层土壤(10~20 cm)含水量无差异(图4a)。狼毒斑块内表层土壤全氮、硝态氮和铵态氮含量显著高于斑块外(图3c, h, i), 防除狼毒斑块外土壤硝态氮和铵态氮显著高于未防除狼毒斑块外(图3h, i)。防除狼毒斑块内深层土壤全氮含量高于狼毒斑块外, 而未防除狼毒斑块内深层土壤全氮含量显著低于斑块外(图4c)。未防除狼毒斑块内深层土壤硝态氮含量高于斑块外, 而防除狼毒斑块外硝态氮含量显著高于狼毒斑块内(图4h), 未防除狼毒斑块内深层土壤铵态氮含量最高(图4i)。狼毒斑块外土壤全钾含量显著高于斑块内, 而狼毒斑块内0~10 cm土壤速效钾含量显著高于斑块外(图3e和图4e)。狼毒斑块内外土壤全磷含量无差异(图3d和图4d), 狼毒斑块内表层土壤速效磷含量显著高于狼毒斑块外(图3f), 而狼毒斑块外深层土壤速效磷含量显著高于狼毒斑块内(图4f)。未防除狼毒斑块内表层土壤有机质含量显著高于狼毒斑块外(图3j), 而防除狼毒斑块外深层土壤有机质含量显著高于防除狼毒斑块内(图4j)。
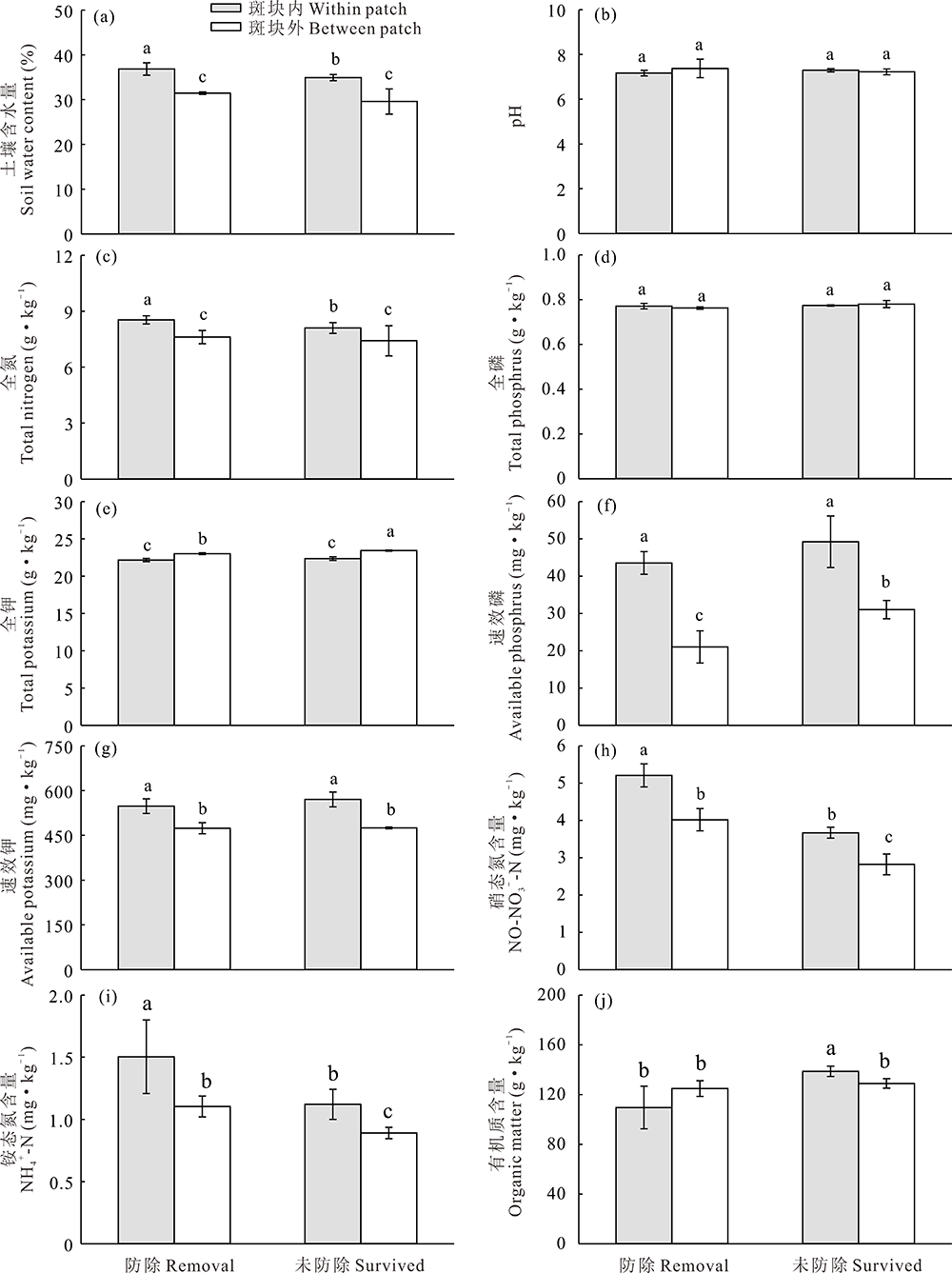 | 图3 围封条件下未防除和防除狼毒0~10 cm土层狼毒斑块内外土壤理化性质变化Fig.3 0-10 cm layer soil physical-chemical properties of natural surviving and removing treatment within or between patch of S. chamaejasme in enclosed plot |
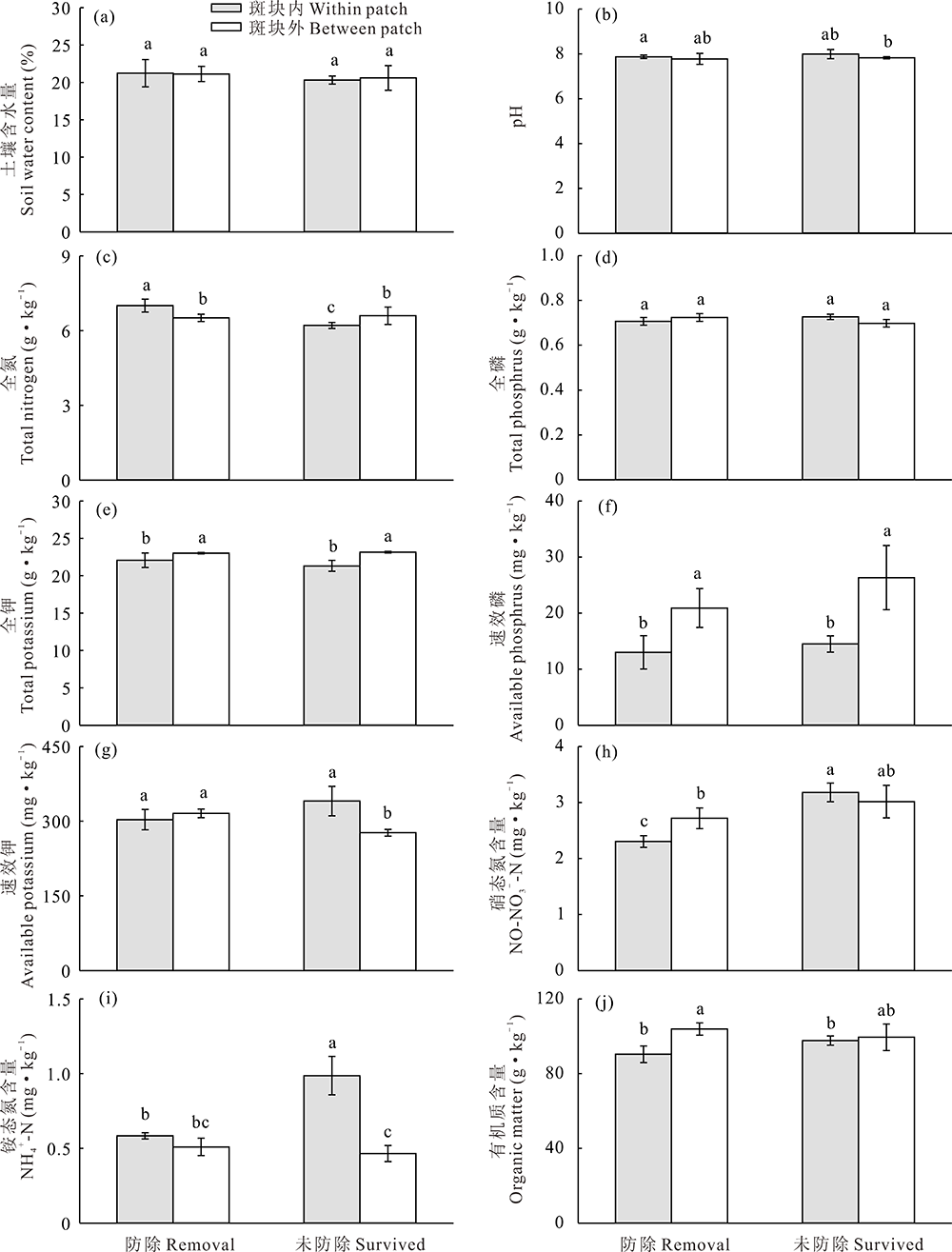 | 图4 围封条件下未防除和防除狼毒10~20 cm土层狼毒斑块内外土壤理化性质变化Fig.4 10-20 cm layer soil physical-chemical properties of natural surviving and removing treatment within or between patch of S. chamaejasme in enclosed plot |
草地围封条件下, 防除狼毒能显著影响狼毒斑块内外土壤微生物量碳、氮和磷含量(图5)。狼毒斑块外土壤微生物量碳显著高于防除狼毒斑块内; 其中, 未防除狼毒斑块外土壤微生物量碳含量最高(图5a)。同时, 防除狼毒斑块外土壤微生物量氮也显著高于防除狼毒斑块内, 而未防除狼毒斑块内土壤微生物量氮显著高于狼毒斑块外(图5b)。另外, 防除狼毒斑块内土壤微生物量磷却显著高于防除狼毒斑块外, 而未防除狼毒斑块外土壤微生物量磷显著高于狼毒斑块内(图5c)。
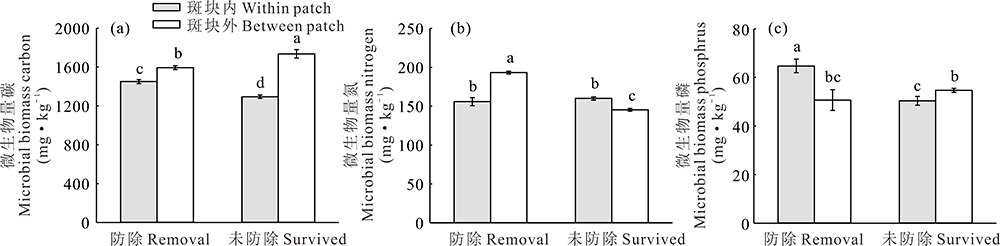 | 图5 围封条件下未防除和防除狼毒斑块内外土壤微生物量变化Fig.5 Soil microbial biomass of natural surviving and removing treatment on within or between patch of S. chamaejasme in enclosed plot |
众多研究表明, 毒害草对同域其他植物充当“ 生态庇护所” 作用, 不仅降低草食家畜对优良牧草的采食风险, 而且为耐牧性差的植物提供生长和繁殖契机; 另外, 毒害草不但调节草地植物个体的空间分布格局, 而且影响整个草地群落结构和生态系统稳定性[29, 30, 31]。狼毒是高寒草原退化阶段主要滋生毒害草种类, 整株含有黄酮类、香豆素类、木酚素、双萜类、倍半萜烯、苯丙醇苷类、挥发油等化学物质[4, 32], 这些化学物质通过雨水淋溶和气体挥发方式直接影响草食家畜对狼毒周围植物的采食行为, 降低草食家畜对狼毒斑块内植物的采食频次, 导致狼毒斑块内植物种类和生物量显著高于斑块间[14]。本研究通过1年围栏封育后, 围封区狼毒斑块内生物量和多样性指数显著高于斑块间(图1和图2), 说明狼毒型退化草地经历长期高强度家畜采食后, 短期禁牧虽能消除草食家畜对优良牧草过度啃食的现象; 而并没有为优良牧草创造快速恢复生长并成为草地优势种的契机; 从侧面反映短期围封并未缩减狼毒的生态位宽度, 且出现狼毒斑块内物种丰富和生物量积累较多的现状, 这与赵成章等[8]研究发现草地围封2年后, 狼毒型天然退化草地禾本科植物生长缓慢的研究结果相似。前期研究发现, 喷施“ 狼毒净” 和“ 灭狼毒” 防除狼毒1年后, 禾本科牧草产量快速增加, 说明使用除草剂能短期内打破狼毒与同域植物间的竞争绝对优势, 导致植物群落结构发生重组和植物多样性发生变化[7, 8, 33]。本研究通过调查狼毒斑块内外植物生物量和物种组成, 发现狼毒防除后狼毒斑块内植物生物量和物种多样性快速降低(图1和图2), 这一研究结果为采用化学除草剂防除狼毒后可食牧草比例和多样性水平提高提供直接证据。另外, 鲍根生等[14]研究发现天然草地持续放牧条件下, 防除狼毒后狼毒斑块内生物量显著高于狼毒斑块外; 而本研究发现围封条件下防除狼毒斑块内生物量却显著低于斑块外; 说明高强度放牧条件下狼毒防除可能造成斑块内植物丧失“ 生态庇护所” , 导致斑块内植物生物量和物种多样性降低; 去除草食家畜的直接影响(草地围封), 防除狼毒可为斑块外植物创造生长和繁殖契机, 导致斑块外植物快速生长。由此可见, 草地围封条件下防除狼毒能短期内加速禾本科等可食牧草的生长, 使狼毒型退化草地逐渐向以禾草为优势种的高寒草原演替, 成为狼毒大面积发生草地治理的有效治理措施[21, 22, 23]。
土壤作为植物生存和生长的物质载体, 成为草地生态系统进行物质和能量交换的重要场所, 同时也是植物、土壤动物和土壤微生物获取生存资源的重要媒介[34]。天然草地从原生阶段向重度退化阶段演替过程中, 土壤物理结构、养分、微生物种类和多样性水平也呈现板结、贫瘠和微生物活动衰退等土壤退化特征[35]。植物种类变化是土壤退化过程最直观的表现, 植物— 土壤界面对草地退化过程表现出明显负效应, 导致重度退化草地阶段呈现以狼毒等毒害草为草地优势种的退化景观[36, 37]。本研究发现狼毒型退化草地围封1年后, 狼毒斑块内土壤有机质含量显著高于斑块外(图3j), 造成斑块内有机质含量增加的原因为:1)狼毒斑块内植物凋落物产量较高; 2)狼毒根系分泌酚类、黄酮类等化学物质影响斑块内植物根际周围活动的土壤节肢动物数量和活动能力降低, 造成根际周围未分解凋落物增加; 而狼毒防除后斑块内外土壤有机质含量差异不显著(图3j), 这与狼毒防除后斑块内凋落物数量降低、狼毒根部腐烂和斑块内土壤动物活动频繁等因素有关[1, 14]。植物地上茎叶凋落物和根系残存物成为影响土壤微生物量和活性的关键因素, 因为根系周围碳水化合物、羧酸类和氨基酸等物质主要来源于植物凋落物和根系残存物, 而这些物质是土壤微生物生存所需的必需物质[38]。
围封条件下, 未防除和施用除草剂狼毒斑块内表层土壤速效磷含量高于斑块间(图3f和图4f), 而深层狼毒斑块内土壤速效磷含量却低于斑块外(图 4f); 同时, 不同土层深度全磷含量差异不显著(图3d和图4d)。这与鲍根生等[14]天然草地在持续放牧情况下, 未防除和防除狼毒斑块内外磷元素变化趋势一致。然而, Sun等[15]在松潘天然草原研究结果却发现狼毒斑块外速效磷和全磷含量高于斑块间; 造成这种差异的原因可能与青藏高原和若尔盖草原地区土壤理化性质、土壤磷元素含量、植被类型、气候因子和水热条件等因素存在差异有关[14, 15, 39, 40]。另外, 本研究发现防除狼毒斑块内土壤微生物磷含量高于狼毒斑块间, 而未防除狼毒斑块内微生物量磷却低于狼毒斑块间(图5c)。说明短期围封条件下防除狼毒造成狼毒斑块内微生物量磷含量降低, 而狼毒斑块间参与土壤微生物磷循环和代谢微生物种类和数量增加, 这与施用“ 狼毒净” 降低斑块土壤微生物活性和狼毒根系腐烂影响有关磷元素运转和吸收微生物种类和数量有关[41]。
狼毒型退化草地中狼毒斑块表现出明显氮素富集效应[14, 15]。短期围封条件下, 狼毒斑块内全氮、速效氮和铵态氮含量呈现氮素蓄积现象, 说明短期围封并未影响狼毒对土壤中总氮素摄取和速效氮的循环和转化能力。然而, 防除狼毒能短期内消除狼毒对氮素占有能力, 驱动氮素向狼毒斑块间转移, 导致防除狼毒斑块间氮素含量高于狼毒斑块间。同时, 本研究发现防除狼毒斑块间微生物量氮含量显著低于狼毒斑块内, 而未防除狼毒斑块内微生物量氮含量显著高于狼毒斑块间(图5b), 这一研究结果与Sun等[15]在松潘狼毒型退化草地研究结果相同, 而与Shi等[42]黄帚橐吾(Ligularia virgaurea)斑块内微生物量氮含量显著低于斑块外研究结果相悖。可见, 狼毒和黄帚橐吾在退化草地中形成斑块过程对氮素利用方式和转化效率存在明显差异, 因为狼毒只具有明显粗壮轴根对狼毒斑块内氮素利用效率低[15], 而黄帚橐吾兼备粗壮的须根根系和发达的横走根状茎造成斑块内氮素消耗速率高处于亏空状态[42, 43]。
综上所述, 狼毒型退化草地采用短期围封并不能实现可食牧草产量提升的目标, 长期(2~5年)围封可进一步恶化狼毒生存环境, 从而为禾本科种群扩张和稳定提供契机, 并最终形成以禾草为优势种的草地群落[15]。另外, 采用“ 狼毒净” 除草剂能短期消除狼毒在草地群落物种间的竞争优势[7, 14], 辅以草地围封措施却能短期内打破狼毒斑块对草地土壤养分的涵养功能, 导致草地土壤养分循环过程和元素代谢途径发生变化, 而这些过程也进一步影响草地微生物量和微生物活性, 反馈至地上植被界面并驱动植物群落结构和物种组成发生变化。可见, 针对狼毒型退化草地需采用“ 狼毒净” 化学除草剂快速降低狼毒种群并辅以长期围封(禁牧)可实现可食牧草比例提升和草地可持续利用的目标。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|