
{kind=link}
{kind=link}
{kind=link}
中国蓼科植物花被片和果实形态结构及其系统学价值
[王靖茹 , 刘玫
, 刘玫* , 茹剑, 曹董玲, 程薪宇, 张欣欣]
, 刘玫, 茹剑, 曹董玲, 程薪宇, 张欣欣]
|
|


作者简介:王靖茹(1989-),女,黑龙江鹤岗人,在读硕士。E-mail:wangjingrugd@126.com
首次采用了外部特征的观察,果皮细胞分离及GMA半薄切片法,深入研究了中国蓼科9属、64种、2变种的花被片及果实的形态结构。结果显示花被片边缘全缘,微波状或锯齿状。三出脉,网状脉或羽状脉。脉序的特征在属内是稳定的。果实多为三棱,双凸镜和近球形。外果皮几乎全为石细胞,石细胞多为镶嵌或栅栏状排列,具7种类型:1)近椭圆形,细胞壁波状;2)长柱状,细胞壁波状弯曲;3)近长方形,细胞壁具突起;4)长圆柱形,细胞壁具突起;5)近长方形,细胞壁平直;6)近椭圆形,细胞壁具突起;7)长条形,细胞壁波状。蓼亚科具类型1~5,酸模亚科具类型6和7。外果皮和中果皮细胞有单宁。本研究为中国蓼科的分类提供花被片及果实结构的依据,为分子系统学揭示的蓼科不是一个自然的类群提供了形态学支持,并为进一步深入研究蓼科的系统学奠定了形态学基础。
The structures of tepals and fruits of 64 species and 2 varieties representing 9 genera of Polygonaceae in China were studied using glycol methacrylate (GMA) section, peeling and separation of the exocarp. Features recorded included whether tepal margins were entire, undulate, or serrate and whether veins were trinervious, netlike or pinnate, although only one venation pattern was observed among species of any one genus. In most cases fruit shape was trigonous, biconvex, or nearly-spherical. In most of taxa, the exocarp consisted of stone cells which were usually arranged in an inlaid or paliform pattern. There were 7 types of stone cells: nearly elliptical with undulate wall, elongate cylindrical with undulate wall, nearly rectangular with protuberant wall, elongate cylindrical with protuberant wall, nearly rectangular with straight wall, nearly elliptical with protuberant wall, and elongate with undulate wall. The first five types were found in the subfamily Polygonideae and the sixth and seventh types in the subfamily Rumicoideae. Tannins occurred in the exocarp and mesocarp of the fruits. This study provides morphological information ontepals and fruit structure in the Polygonaceae, and from a taxonomic perspective supports the conclusion from molecular study that Polygonaceae is not a natural group.
蓼科(Polygonaceae)世界约50余属, 1150余种。我国有13属, 235种, 37变种, 全国各地均有分布[1], 其中一些类群, 如大黄属(Rheum)及何首乌属(Fallopia)具清热解毒及消炎等功效[2, 3, 4]。
营养器官的形态特征通常为被子植物的分类依据[5, 6, 7]。蓼科依据花被片的特征被分为两个亚科:蓼亚科(Polygonideae), 除了木蓼属(Atraphaxis)外, 花被片均排列为一轮; 酸模亚科(Rumicoideae)的花被片排列为两轮[1]。中国蓼族的花被片具羽状脉, 三出脉和单一脉[8]。侯元同等[9]观察到萹蓄组(Sect. Avicularia)的果实分长果及短果。曲畅游[10]指出蓼科果皮平滑, 或具瘤状颗粒及各种纹饰。还有一些学者注意到蓼科的果皮结构, 如张晓霞等[11]指出山东蓼属(Polygonum)的外果皮细胞为方形, 短矩形或狭长矩形, 垂周壁平直或波纹状, 细胞腔具树状分枝或二叉分枝。李淑久等[12]描述荞麦属(Fagopyrum)的外果皮由厚壁组织组成。
近年来, 分子系统学的研究揭示蓼族或广义蓼属(包括蓼族下的类群)不是自然的类群。孙伟[13]指出蓼属拳参组(Sect. Bistorta)应独立成属, 春蓼组(Sect. Persicaria)、头状蓼组(Sect. Cephalophilon)、刺蓼组(Sect. Echinocaulon)和金线草组(Sect. Tovara)合并为春蓼属(Persicaria)。然而闵运江[14]指出蓼属中的春蓼组, 刺蓼组, 拳参组, 分叉蓼组(Sect. Aconogonon)应分别独立成属, 其中分叉蓼组中的西伯利亚蓼(Polygonum sibiricum)应提升为西伯利亚蓼属(Knorringia)。
由于缺少对蓼科植物花被和果实结构的系统研究, 蓼科不同类群的划分没有详细的花被和果实结构的形态学依据。已有的分子系统学研究缺少形态学支持。本文深入研究了中国蓼科9属、64种、2变种(分别属于2亚科及3个族)花被及果实的微形态结构, 旨在完善中国蓼科花被和果实形态学, 为该科的分类及分子系统学研究提供形态学依据。
实验材料为中国蓼科9属, 64种, 2变种的果实, 其中蓼亚科6属49种, 2变种, 酸模亚科3属15种。2012及2013年采自野外的植物经王臣教授鉴定, 标本存放于哈尔滨师范大学标本室(HANU), 部分材料取自于东北林业大学植物标本馆(NFFI)、东北农业大学植物标本馆(NEAU)、沈阳生态研究所植物标本馆(IFP)以及华南植物园标本馆(IBSC)。物种名称及凭证标本详尽信息见表1。
| 表1 中国蓼科所研究的物种、凭证标本及采集地 Table 1 Polygonaceae species in China used for the study, together with voucher specimen details and locality |
花被、果实形态及果皮结构的观察:在解剖镜(Olympus SZX 16)下观察花被片和果实形态并测量果实大小及花被片长度, 之后将花被片(每物种取2片)及果实(每物种取2个)放入热水(约80℃), 浸泡约2 h, 待其吸水膨胀后取出并放入次氯酸钠脱色[15], 待材料脱色后取出并水洗, 在Olympus BX53显微镜下观察花被片脉序类型。果实及花被均用Olympus DP 26成像系统照相。撕取经吸水膨胀并脱色后果实的果皮, 将其置于载玻片, 加入1~2滴50%甘油水溶液, 盖上盖玻片, 轻敲盖片, 使果皮的细胞彼此分离。用具Olympus DP 26成像系统的显微镜(Olympus BX53)观察细胞的形态并照相。此外, 用亚铁盐鉴定细胞内似单宁物质[16]。
GMA(glycol methacrylate, 乙二醇甲基丙烯酸酯)半薄切片:吸水膨胀后的果实(每物种取2个)经FAA(formalin-acetic acid-alcohol, 福尔马林-醋酸-酒精)固定不少于24 h, 根据Feder和O’ Brien[17]的方法, 固定后的材料经50%酒精(2次, 每次4~6 h), 100%酒精(2次, 每次4~6 h), 异丙醇(2次, 每次6 h)及正丁醇(2次, 每次6 h)脱水, 之后进入GMA渗透3次。第1和2次各为1 d, 第3次不少于5 d, 之后将装有材料和GMA的胶囊置于60℃温箱24 h。用Leica Ultralcut R切片机切片, 厚度为2~3 μ m。同样按照Feder和O’ Brien[17]的方法, 将切片用希夫试剂-考马斯亮蓝染色, 中性树胶封片, 用具Olympus DP 26成像系统的显微镜(Olympus BX53)观察并照相。
本文深入研究了中国蓼科9属64种2变种花被片及果实的形态结构, 揭示了花被片的形状, 脉序, 果皮(外果皮, 中果皮及内果皮)的微形态结构, 详细特征见表2及图1~图3。
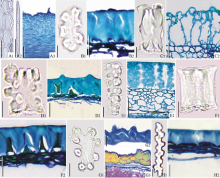
所研究的蓼科物种, 花被片长度变化较大, 蓼属花被片较小, 篱蓼较大, 变化范围为1.5~18.0 mm。花被片分4种形状:椭圆形(如狭叶蓼及酸模叶蓼, 图1B)、卵形(如小果酸模及毛脉酸模, 图1F1)、心形(如狭叶酸模及木蓼, 图1G)和近圆形(如直根酸模及酸模, 图1H)。花被片的形状在族内不同属均有变化(表2)。花被片边缘为全缘(如酸模叶蓼及两栖蓼, 图1D1)、微波状(如直根酸模及刺木蓼, 图1H)或锯齿状(如狭叶酸模及羊蹄, 图1G)。酸模族花被片边缘具3种类型, 木蓼族木蓼属微波状, 沙拐枣属全缘, 蓼族全缘或锯齿状。何首乌属(蓼族)花被片具翅(如齿翅蓼及篱蓼, 图1A)。酸模属(酸模族)的长刺酸模具1对刺(图1I), 刺酸模及黑龙江酸模具2~3对刺(图1E), 酸模属的一些物种花被片背面还具小瘤(如刺酸模及小果酸模, 图1E)。花被片脉序分为羽状脉(如拳参及西伯利亚蓼, 图1C)、网状脉(如直根酸模及小果酸模, 图1H, 图1F2)及三出脉, 三出脉为花被片的基部伸出3条大小相似的脉, 每条脉有小的分枝或分枝不明显(如狭叶蓼及珠芽蓼, 图1B), 少数物种3条脉的末端, 即在花被片边缘具锚状分枝(如酸模叶蓼, 图1D1)。酸模族具3种脉序, 蓼族具羽状脉或三出脉, 木蓼族具网状脉。
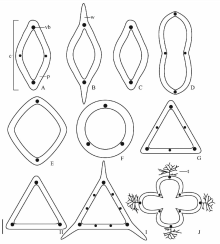
果实为褐色(如金线草及酸模叶蓼, 图1J)或黑色(如柳叶刺蓼及长鬃蓼, 图1M), 长1.1~14.0 mm, 宽0.6~11.0 mm。除了沙拐枣属果实具4心皮外, 每一个族果实心皮数目为2或3, 其中多数为3心皮, 如蓼族的何首乌属及荞麦属具3心皮, 金线草属具2心皮。3心皮的果实多数为三棱形, 中部或中下部膨大, 上部渐尖(如巴天酸模及阿扬蓼, 图1P), 其横切面为三角形, 棱脊为2心皮愈合处(如谷地蓼及卷茎蓼, 图3G), 部分3心皮的果实近球形(如杠板归及稀花蓼, 图1N), 其横切面为圆形(如稀花蓼, 图3F)。2个心皮的果实为卵圆双凸形, 横切面椭圆形, 心皮的背缝线处微突起(如金线草, 图1J, 图3A); 近圆双凸形(如柳叶刺蓼及两栖蓼, 图1M), 横切面椭圆形, 心皮的背缝线处突起(如尼泊尔蓼, 图3E); 近圆双凹形, 横切面椭圆形, 心皮的背缝线处凹陷(如酸模叶蓼, 图1D2, 图3D)。沙拐枣果实椭圆形(包括刺), 横切面呈十字形, 心皮的背缝线处隆起, 生有分枝状刺, 2心皮愈合处凹陷(图1K1, 图1K2, 图3J)。山蓼属和大黄属的果实具2个或3个翅, 沙拐枣果实每条果肋上有2或3个分枝状刺, 翅和刺由外果皮和中果皮向外延伸形成(如山蓼, 药用大黄, 滇边大黄, 沙拐枣, 图1L, 图1O, 图1K1, 图3B, 图3I, 图3J)。
| 表2 中国蓼科花被片和果实的形态结构 Table 2 Morphology and structure of tepal and fruit of Polygoaceae in China |
 | 图1 中国蓼科植物花被片和果实 示花被片及果实的形态。 A.齿翅蓼花被片; B.狭叶蓼花被片; C.拳参花被片; D1.酸模叶蓼花被片; D2.酸模叶蓼果实; E.刺酸模花被片; F1~2.小果酸模花被片; G.狭叶酸模花被片; H.直根酸模花被片; I.长刺酸模花被片; J.金线草果实; K1.沙拐枣果实; K2.沙拐枣果实的刺; L.山蓼果实; M.柳叶刺蓼果实; N.杠板归果实; O.药用大黄果实; P.巴天酸模果实。比例尺:图A, B, C, D1~2, E, F1~2, G, H, I, J, L, M, N, P=1 mm; K1~2=3 mm; O=2 mm。Fig.1 The tepals and fruits of Polygonaceae in China Showing the shapes of tepals and fruits. A. Tepal of Fallopia dentato-alata; B. Tepal of Polygonum angustifolium; C. Tepal of Polygonum bistorta; D1. Tepal of Polygonum lapathifolium; D2. Fruit of Polygonum lapathifolium; E. Tepal of Rumex maritimus; F1~2. Tepal of Rumex microcarpus; G. Tepal of Rumex stenophyllus; H. Tepal of Rumex thyrsiflorus; I. Tepal of Rumex trisetifer; J. Fruit of Antenoron filiforme; K1. Fruit of Calligonum mongolicum; K2. Thorn of fruit of Calligonum mongolicum; L. Fruit of Oxyria digyna; M. Fruit of Polygonum bungeanum; N. Fruit of Polygonum perfoliatum; O. Fruit of Rheum officinale; P. Fruit of Rumex patientia. Scale bar=1 mm in A, B, C, D1-2, E, F1-2, G, H, I, J, L, M, N, P; 3 mm in K1-2; 2 mm in O. |
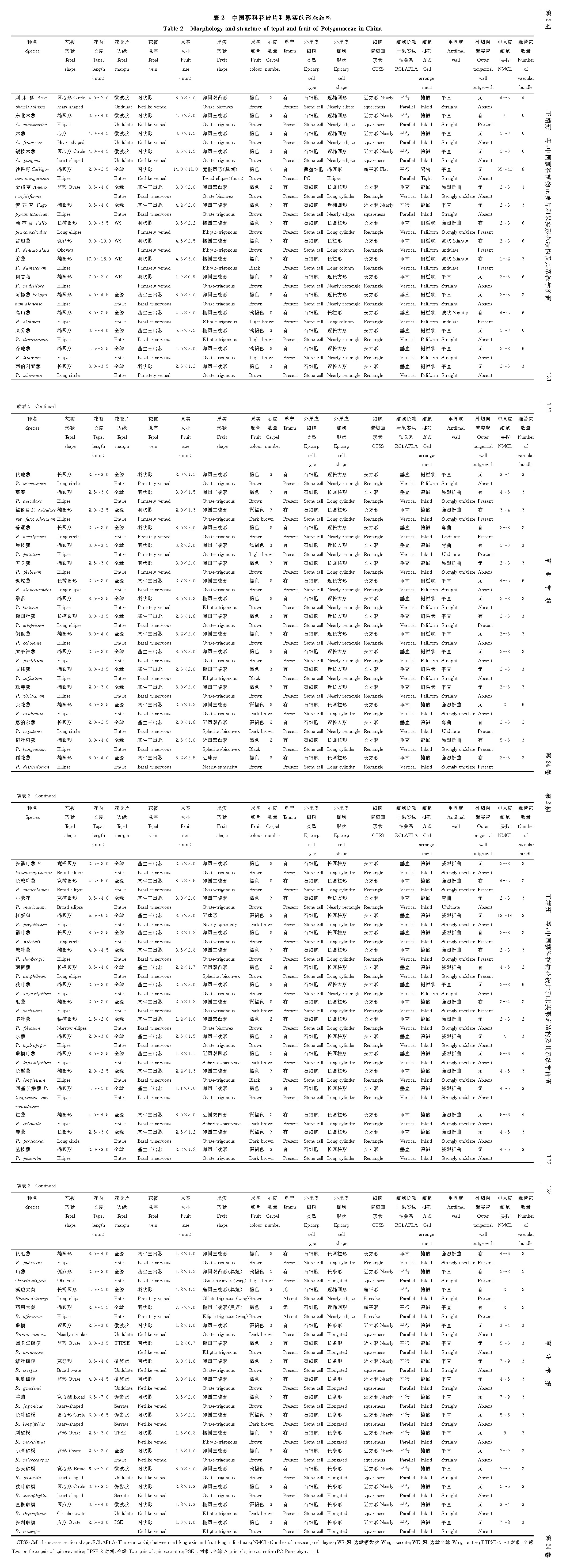
 | 图2 中国蓼科植物果实 示果实结构及石细胞。 A1.沙拐枣薄壁细胞; A2.沙拐枣纤维; A3.沙拐枣果实结构; B1.苦荞麦石细胞; B2.苦荞麦果实结构; C1.篱蓼石细胞; C2.篱蓼果实结构; D1.普通蓼石细胞; D2.普通蓼果实结构; E1.春蓼石细胞; E2.春蓼果实结构; F1.珠芽蓼石细胞; F2.珠芽蓼果实结构; G1.滇边大黄石细胞; G2.滇边大黄外果皮结构; G3.滇边大黄果实结构; H1.直根酸模石细胞; H2.直根酸模果实结构。比例尺:图A1, A3=50 μ m; A2, G3=100 μ m; B1, C1, D1, E1~2, F1, G1=20 μ m; B2, D2, F2=30 μ m; C2, G2, H1~2=10 μ m。Fig.2 Fruits of Polygonaceae in China Showing fruit structures and stone cells. A1. Parenchyma cell of Calligonum mongolicum; A2. Fiber of Calligonum mongolicum; A3. Fruit structure of Calligonum mongolicum; B1. The stone cell of Fagopyrum tataricum; B2. Fruit structure of Fagopyrum tataricum; C1. The stone cell of Fallopia dumetorum; C2. Fruit structure of Fallopia dumetorum; D1. The stone cell of Polygonum humifusum; D2. Fruit structure of Polygonum humifusum; E1. The stone cell of Polygonum persicaria; E2. Fruit structure of Polygonum persicaria; F1. The stone cell of Polygonum viviparum; F2. Fruit structure of Polygonum viviparum; G1. The stone cell of Rheum delavayi; G2. Epicarp structure of Rheum delavayi; G3. Fruit structure of Rheum delavayi; H1. The stone cell of Rumex thyrsiflorus; H2. Fruit structure of Rumex thyrsiflorus. Scale bar=50 μ m in A1, A3; 100 μ m in A2, G3; 20 μ m in B1, C1, D1, E1-2, F1, G1; 30 μ m in B2, D2, F2; 10 μ m in C2, G2, H1-2. |
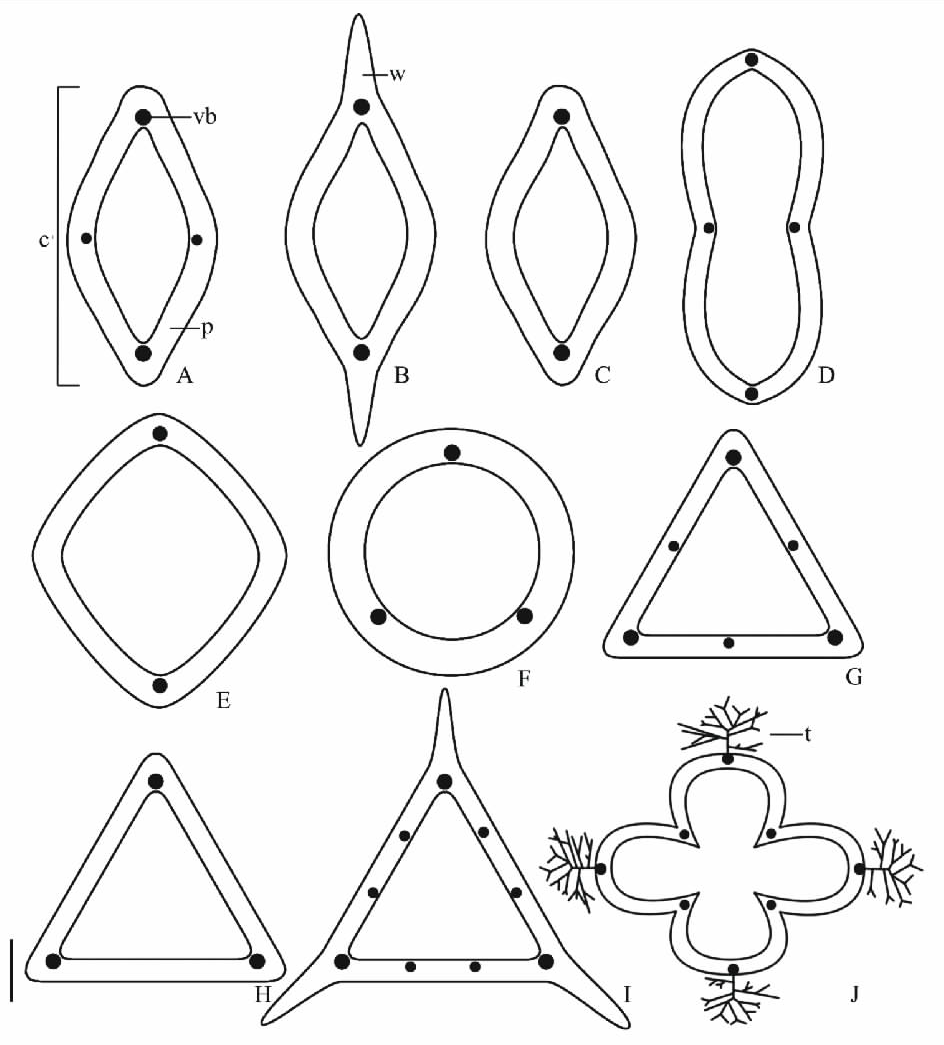 | 图3 中国蓼科植物果实横切面示意图 A.金线草; B.山蓼; C.多叶蓼; D.酸模叶蓼; E.尼泊尔蓼; F.稀花蓼; G.谷地蓼; H.春蓼; I.滇边大黄; J.沙拐枣。缩写:c=心皮; p=果皮; t=刺; vb=维管束; w=翅。比例尺:A~H=1 mm; I=2 mm; J=3 mm。Fig.3 Diagrammatic interpretation of fruits (in cross section) of Polygonaceae in China A. Antenoron filiforme; B. Oxyria digyna; C. Polygonum foliosum; D. Polygonum lapathifolium; E. Polygonum nepalense; F. Polygonum dissitiflorum; G. Polygonum limosum; H. Polygonum persicaria; I. Rheum delavayi; J. Calligonum mongolicum. Abravation:c=carpel; p=pericarp; t=thorn; vb=vascular bundle; w=wing. Scale bar=1 mm in A-H; 2 mm in I; 3 mm in J. |
沙拐枣属的外果皮由一层薄壁细胞组成(如沙拐枣, 图2A1), 而其余的所有物种外果皮由一层石细胞组成, 细胞外壁平滑(如春蓼及直根酸模, 图2E2)或具小突起(如篱蓼及普通蓼, 图2C2)。石细胞具7种类型:1)近椭圆形, 细胞长轴与果实纵轴平行, 细胞壁(除外切向壁)呈波状, 镶嵌排列, 壁厚, 中央空腔小, 横切面近方形(如苦荞麦及锐枝木蓼, 图2B1, 图2B2); 2)长柱形, 细胞长轴与果实纵轴垂直, 栅栏状排列, 细胞壁(除内切向壁)波状弯曲, 壁薄, 中央空腔大, 横切面长方形(如篱蓼及齿翅蓼, 图2C1, 图2C2); 3)近长方形, 细胞长轴与果实纵轴垂直, 细胞壁(除内切向壁)具少数不规则突起, 镶嵌排列, 壁厚, 腔较大, 横切面长方形(如普通蓼及尼泊尔蓼, 图2D1, 图2D2); 4)长圆柱形, 细胞长轴与果实纵轴垂直, 细胞壁(除内、外切向壁)具许多不规则突起, 镶嵌排列, 壁薄, 腔较大, 横切面长方形(如春蓼及金线草, 图2E1, 图2E2); 5)近长方形, 细胞长轴与果实纵轴垂直, 细胞壁平直, 栅栏状排列, 壁厚, 腔较小, 横切面长方形(如珠芽蓼及倒根蓼, 图2F1, 图2F2); 6)近椭圆形, 细胞扁平, 细胞长轴与果实纵轴平行, 细胞壁外壁四周具突起, 镶嵌排列, 壁厚, 腔小, 横切面扁平形(如滇边大黄及药用大黄, 图2G1, 图2G2); 7)长条形, 长轴与果实纵轴平行, 细胞壁(除内、外切向壁)呈波状, 镶嵌排列, 壁厚, 腔小, 横切面近方形(如直根酸模及皱叶酸模, 图2H1, 图2H2)。木蓼族具类型1, 蓼族具类型1~5, 酸模族具类型6和7。
沙拐枣属的中果皮由纤维组成(图2A2), 靠近外果皮的纤维与果实纵轴平行, 靠近内果皮的纤维多与果实纵轴垂直(图2A3)。其余所有物种的中果皮由多层薄壁细胞组成, 细胞扁平(如苦荞麦及珠芽蓼, 图2B2)、圆形或近圆形(如篱蓼及春蓼, 图2C2)或椭圆形(如滇边大黄及药用大黄, 图2G3)。中果皮维管束为2, 3, 4, 6, 8, 9。所有果实心皮的愈合处均具有1个维管束(如多叶蓼及春蓼, 图3C, 图3H)。沙拐枣属及部分具2心皮或3心皮的果实, 每个心皮中央还具1个(如金线草及谷地蓼, 图3A, 图3G)。大黄属3心皮果实, 每个心皮中央具2个(如滇边大黄, 图3I)。沙拐枣的维管束延伸至刺的基部(图3J)。内果皮常为一层扁平的薄壁细胞。除大黄属外, 蓼科果实的外果皮和中果皮中有单宁。
1. 花被具翅何首乌属 Fallopia
1. 花被无翅2
2. 果实具翅或刺3
2. 果实无翅或刺5
3. 果实具翅, 外果皮具石细胞4
3. 果实具刺, 中果皮具纤维沙拐枣属Calligonum
4. 果实为椭圆三棱形大黄属 Rheum
4. 果实为卵圆双凸形山蓼属 Oxyria
5. 外果皮石细胞垂周壁平直蓼属 Polygonum
5. 外果皮石细胞垂周壁不平直6
6. 外果皮石细胞垂周壁呈波状7
6. 外果皮石细胞垂周壁具不规则突起10
7. 外果皮石细胞长轴与果实纵轴平行8
7. 外果皮石细胞长轴与果实纵轴垂直蓼属 Polygonum
8. 外果皮石细胞长条形酸模属 Rumex
8. 外果皮石细胞近椭圆形9
9. 心皮中央具维管束木蓼属 Atraphaxis
9. 心皮中央不具维管束荞麦属 Fagopyrum
10. 果实为卵圆双凸形11
10. 果实非卵圆双凸形蓼属 Polygonum
11. 维管束数量为4金线草属 Antenoron
11. 维管束数量为2蓼属 Polygonum
花被片脉序:蓼科花被片脉序通常在属内(如金线草属, 沙拐枣属, 木蓼属, 何首乌属, 荞麦属, 山蓼属, 酸模属和大黄属)是稳定的特征, 可以作为属的分类依据。例如, 木蓼属所研究的物种都具有网状脉, 何首乌属都具有羽状脉。但是在族内不同的属脉序类型是有变化的, 如酸模族山蓼属具三出脉, 大黄属具羽状脉。李安仁[1]将木蓼属及沙拐枣属归于木蓼族。刘明珍[18]指出木蓼属及沙拐枣属茎解剖结构相似, 木质部发达, 皮层内厚角组织发达, 支持李安仁[1]的观点。我们的研究显示木蓼属及沙拐枣属花被片均为网状脉, 但木蓼属的花被片排成2轮, 内轮花被片较大, 包被果实, 边缘微波状。沙拐枣花被片排成1轮, 较小, 果期反折, 不包被果实, 边缘全缘, 其果实特征亦有区别(见下)。何首乌属原位于蓼属蔓蓼组(Sect. Tiniaria)[19], 后来从蓼属分离出来, 作为何首乌属[1, 20, 21]。本研究揭示何首乌属植物花被片具翅, 而蓼属其他类群花被片无翅, 支持建立何首乌属。此外, 我们观察到蓼属不同类群花被片特征不同, 如萹蓄组具羽状脉, 蓼组、头状蓼组和刺蓼组具基生三出脉, 支持闵运江[14]的观点, 即蓼族不是个自然的类群。早期经典分类学研究及分子系统学将西伯利亚蓼从蓼属的分叉蓼组分出, 作为西伯利亚蓼属[22]。花被脉序为羽状脉的结构显示西伯利亚蓼及其他分叉蓼组花被片的脉序不同, 前者为羽状脉, 后者为基生三出脉, 支持西伯利亚蓼独立为属[23]。
果实翅、刺、心皮数目及果皮结构:翅及果实心皮的数目在多数类群是稳定的性状, 酸模亚科的山蓼属及大黄属果实均具翅。金线草属及山蓼属所有物种均具2心皮, 何首乌属, 荞麦属, 大黄属, 酸模属均具3心皮(但蓼属和木蓼属具2或3心皮), 沙拐枣属均具4心皮。外果皮结构显示蓼亚科及酸模亚科的石细胞形态不同, 如前者石细胞通常垂直于果实纵轴, 后者通常平行于果实纵轴。蓼亚科, 蓼族的蓼属、何首乌属及金线草属均具有长圆柱形石细胞, 细胞壁不规则突起, 分子系统学揭示出金线草与蓼属亲缘关系很近, 前者不应成为独立的属[24], 但何首乌属与金线草及蓼属的关系需要进一步研究。Linnaeus[25]以Calligonum polygoboides为模式建立沙拐枣属。我们的研究显示沙拐枣属具4心皮, 果肋上具刺, 外果皮由薄壁细胞组成, 中果皮由纤维组成, 明显不同于木蓼属的果实(2或3心皮, 无翅及刺, 外果皮为石细胞), 因而木蓼族是否为自然类群需要进一步研究。Meisner[26]将荞麦属作为蓼属中的一个组, 后将其从蓼属分出作为独立的属[27]。周忠泽等[28]指出荞麦属的花粉形态极为一致, 其外壁纹饰均为细网状, 萌发孔为三孔沟, 支持将荞麦属从蓼属中分出独立成属, 这一研究得到分子系统学的支持[13]。荞麦属果实结构不同于蓼科其他属(即外果皮石细胞近椭圆形, 垂周壁波状弯曲, 维管束为3)同样支持将荞麦属作为单独的一个属。张晓霞等[11]认为头状蓼组和刺蓼组与春蓼组的外果皮细胞腔近狭长, 具树状分枝, 垂周壁强烈折曲或波纹状, 具有很高的相似性, 应将前两个组归于春蓼组, 并将该组提升为春蓼属。分子系统学上的研究, 春蓼组、头状蓼组、刺蓼组和金线草组合并为春蓼属[13], 我们的研究显示这4个组的果实结构极为相似, 外果皮石细胞多为长圆柱形, 垂周壁具不规则突起, 镶嵌排列, 支持张晓霞等[11]的观点, 同时为分子系统学上的研究提供了形态学依据。此外, 蓼属中拳参组与分叉蓼组的外果皮石细胞多为近长方形, 细胞栅栏状排列, 作者认为两组可能有很近的亲缘关系。褐鞘蓼(Polygonum fusco-ochreatum)是Komarov[29]建立的, 李安仁[1]将其作为萹蓄的变种。侯元同等[9]通过研究果实表面纹饰, 认为褐鞘蓼不同于萹蓄, 应恢复褐鞘蓼种级。本研究表明, 二者外果皮石细胞结构相同, 均为长圆柱形, 垂周壁具许多不规则突起, 镶嵌排列, 支持李安仁[1]将褐鞘蓼作为萹蓄的变种。
通过深入研究蓼科花被片及果实的形态结构, 揭示了花被片及果实的结构特征, 特别是花被片的脉序及果皮的结构可以作为亚科、族、属, 甚至种的分类依据。本研究完善了中国蓼科植物的形态学研究, 为蓼科的分类及分子系统学的研究提供形态学依据, 并为进一步深入研究蓼科系统学奠定了形态学基础。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [16] |
|
| [18] |
|
| [20] |
|
| [21] |
|
| [24] |
|
| [28] |
|