

{kind=link}
{kind=link}
东北唇形科(Lamiaceae)果实形态与解剖特征及其分类学价值
[茹剑 , 刘玫
, 刘玫* , 王臣, 程薪宇, 梁莉]
, 刘玫, 王臣, 程薪宇, 梁莉]
|
|


作者简介:茹剑(1987-),男,黑龙江大庆人,在读博士。E-mail:rujian1987@163.com
本文首次采用了GMA半薄切片和厚壁区域细胞分离法,深入研究了东北唇形科52种及5变种(位于4亚科26属)的果实形态结构。结果显示,果实多为三棱形,有的物种果实的棱延伸形成翅。果实表面光滑、具网纹或不同类型的毛状体(包括头状或盾状腺毛,单细胞、单列多细胞或分枝非腺毛)。着生面多为圆形,少为直线形或“V”形。果皮分为外果皮、中果皮、厚壁区域和内果皮。晶体常存在厚壁区域。本研究完善了东北唇形科果实形态学,并为进一步研究中国唇形科果实结构和分类学以及分子系统学奠定了基础。
The fruit structures of 52 species and 5 varieties representing 26 genera from 4 subfamilies of Lamiaceae in northeasten China were deeply studied in the paper for the first time, using glycol methacrylate(GMA) sections and cells of the sclerenchyma region were separated for identifying the structure. The results show that the fruits of the family are mainly trigonous in shape, sometimes the edges of the fruits extend and form wing. The fruit surfaces are glabrous, reticulate or with hairs (including capitate or peltate glandular hairs, unicellular, uniseriate multicellular or branched non-glandular hairs). The areola of the fruit is mainly round, barely linear or V-shape. The pericarp could be divided into four parts: exocarp, mesocarp, sclerenchyma region and endocarp. The crystal exists in the sclerenchymatous region. The study completes the fruit anatomy of Lamiaceae in northeasten China and provides the basis for the further research of the fruit anatomy and taxonomy, and molecular systematics of Lamiaceae in China.
唇形科(Lamiaceae)为被子植物较大的科, 分为10个亚科, 约220余属, 3500余种, 我国有7亚科, 即筋骨草亚科(Ajugoideae)、保亭花亚科(Wenchengioideae)、锥花亚科(Prasioideae)、黄芩亚科(Scutellarioideae)、薰衣草亚科(Lavanduloideae)、野芝麻亚科(Lamioideae)和罗勒亚科(Ocimoideae), 99属, 800余种[1]。东北产28属, 74种, 10变种[2]。植物常为多年至一年生草本, 少为半灌木或灌木, 富含芳香油, 具有较高的药用及观赏价值[1, 2, 3, 4]。
唇形科果实, 特别是果皮结构对属和种的分类具有重要作用[5, 6, 7, 8, 9, 10, 11, 12, 13]。Ryding[14, 15, 16, 17, 18]指出唇形科(如罗勒族Ocimeae和薄荷族Mentheae等)的果皮包括4层:外果皮, 中果皮, 厚壁区域及内果皮, 并描述了厚壁区域细胞的类型及晶体的分布。唇形科果实遇水后产生粘液[19, 20]。Budantsev和Lobova[21]依据花序类型、果实形状及表面纹饰等特征将荆芥族(Nepeteae)分为3个类群:类群1包括6属:藿香属(Agastache), Drepanocaryum, Hymenocrater, 扭藿香属(Lophanthus), Marmoritis和荆芥属(Nepeta); 类群2包括3属:Cedronella, 青兰属(Dracocephalum)和裂叶荆芥属(Schizonepeta); 类群3包括2属:活血丹属(Glechoma)和龙头草属(Meehania)。Oran[6]和Habibvash等[8]注意到鼠尾草属(Salvia)果皮的厚度可用于种的分类。地笋属(Lycopus)果皮中具典型适应水媒传播的气室[22]。Kaya和Dï rmencï [9]依据果实表面特征将土耳其荆芥属39种分为3种类型:平滑, 部分平滑及具纹饰。果实特征还被用于区别3种鼠尾草(Salvia hedgeana, S. huberi和S. rosifolia)[23]。近些年随着分子生物学的发展, 人们也将其用于唇形科的系统学研究, 例如Bendiksby等[24]通过核(NRPA2, 5S-NTS)和叶绿体(matK, psbA-trnH, rps16, trnL, trnL-F, trnS-G)DNA序列对野芝麻属(Lamium)进行分析, 强调了系统发育关系及异源多倍体物种的形成。
营养器官(如叶柄和茎)的特征常被作为植物分类的主要依据[25, 26, 27]。然而, 果实的结构常被忽略。唇形科少数已有果实表面特征的报道也仅局限于某个属或少数物种, 例如, 胡珊梅和范崔生[28]揭示了石荠苧属(Mosla)不同物种果实表面特征(包括网纹密度, 网脊形态及网脊内突起)有区别。林文群等[29]指出果实的形状、颜色和表面纹饰是区别唇形科植物的重要依据。邵邻相等[30]观察到3种香茶菜, 香茶菜(Isodon amethystoides), 大萼香茶菜(I. macrocalyx)和显脉香茶菜(I. nervosa)果实表面腺毛和非腺毛数量不同。郑宝江等[31]曾研究了东北地区11属12种果皮的超微结构, 将果皮纹饰分为6种类型(网状、鸟巢状、丝状、丘状、条状和复合纹饰)。迄今为止, 尚无关于我国唇形科果实结构的报道。本研究的目的是通过深入研究东北唇形科26属, 52种及5变种果实的形态结构, 完善东北唇形科果实形态学, 为唇形科的分类及系统学研究提供果实的形态学依据。为进一步研究唇形科分子系统学提供形态学资料。
研究材料为我国东北产唇形科26属, 52种, 5变种的成熟果实, 材料于2013年取自中国科学院沈阳应用生态研究所(Institute of Applied Ecology, Chinese Academy of Sciences, IFP)。物种名称、凭证标本及采集地详细信息见表1, 物种在中国植物志中的分类地位见表2。
观察果实外形及果皮结构特征:在Olympus SZX16解剖镜下观察果实外形, 测量果实大小, 并用Olympus DP26成像系统照相, 此外观察粘液质的形成。将果实水煮约1 h后冷却, 待果实充分吸水膨胀后, 撕取外果皮, 将其放在载玻片上, 加1~2滴50%甘油水溶液, 盖上盖玻片。将其余部分浸泡于NaClO[17], 待材料变软后取出, 水洗后置于载玻片, 加1~2滴50%甘油水溶液, 盖上盖玻片, 轻敲盖片, 使厚壁区域的细胞均匀分散。用Olympus BX51显微镜观察毛状体的类型及厚壁区域细胞特征, 并用Olympus DP70成像系统照相。
GMA(glycol methacrylate, 乙二醇甲基丙烯酸酯)半薄切片:每个物种取2或3个果实, 经吸水膨胀后放入FAA(formalin-acetic acid-alcohol mixture, 福尔马林-醋酸-酒精混合液)固定不少于24 h, 根据Feder和O’ Brien[32]的方法, 将材料浸入GMA中, 前2次渗透时间均不少于24 h, 第3次渗透不少于5 d。将装有材料和GMA的胶囊置于60℃温箱24 h。用Leica Ultralcut R切片机切片, 厚度为1.5~2.0 μ m。同样按照Feder和O’ Brien[32]的方法, 将切片用希夫试剂-甲苯胺蓝染色, 中性树胶封片。在Olympus BX51显微镜下观察, 并用Olympus DP70成像系统照相。
| 表1 东北唇形科果实材料、凭证标本及采集地 Table 1 Taxa of Lamiaceae in northeasten China examined for fruit structures, together with voucher specimen detail and locality |
| 表2 东北唇形科物种的分类地位(中国植物志) Table 2 The taxonomic status of Lamiaceae in northeasten China(Flora of China) |
本文详细研究了东北唇形科26属, 52种及5变种果实的外形及毛状体特征, 果皮结构(包括外果皮, 中果皮, 厚壁区域及内果皮特征)和内含物(图1, 2), 详细特征见表3。
| 表3 东北唇形科果实的形态结构 Table 3 Fruit shape and structure of Lamiaceae in Northeast China |
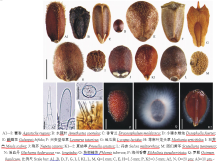
成熟果实为褐色、深褐色或黑色。长0.5~6.0 mm, 宽0.5~2.9 mm。果实常为三棱形, 卵形、椭圆形或近圆球形。三棱形的果实远轴面平或微突, 近轴面中央突起形成脊(如藿香, 图1A1), 顶端圆形(如鼬瓣花, 图1E)或平截(如兴安益母草, 图1F)。水棘针腹面一侧具一条狭长的裂隙, 两侧具短翼(图1B)。地瓜苗的果皮延伸形成翅, 顶端边缘形成锐齿, 远轴面为马蹄状衣领形, 基部不愈合(图1G)。果实着生面呈圆形(如藿香和丹参, 图1A1, L)、三角形(如荨麻叶龙头草, 图1H)、“ V” 形(如香青兰, 图1C)或直线形(如荆芥, 图1J)。果实表面光滑有光泽(如夏枯草, 图1K1)或不光滑, 即具大理石花纹(如鼬瓣花, 图1E)、网状纹饰(如石荠苎, 图1I)、突起(如图们黄芩, 图1M)或具毛状体。毛状体包括非腺毛和腺毛, 多集中于果实顶端(如藿香和兴安益母草, 图1A1, F)和腹面(如石荠苎, 图1I), 少数密被整个果实(如荨麻叶龙头草, 图1H)。非腺毛为单列多细胞(如藿香, 图1A2)或单细胞(如藿香, 图1A3)。腺毛通常具极短的柄(如活血丹, 图1N)。腺毛顶细胞通常为头状, 极少数为盾状(如荠苎属)。块根糙苏具分枝非腺毛, 非腺毛中央有一个具长柄的头状腺毛(图1O)。
部分物种的果实遇水后产生明显的粘液, 粘液乳白色, 呈放射的丝状包裹果实(如罗勒, 图1Q)或为半透明的粘液质(如夏枯草, 图1K2), 一些物种可见粘液填充于外果皮细胞内(如海州香薷, 图1P)。
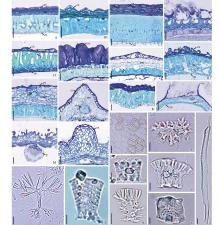
外果皮:由一层细胞组成, 常覆盖角质层。细胞呈柱状(如藿香, 图2A)、扁平长方形(如图们黄芩, 图2N)或外切向壁形成突起(如水棘针和光萼青兰, 图2B1, C1)。部分物种的外切向壁加厚(图2C1, J1)。鼬瓣花的外果皮细胞壁常有网状加厚(图2E1~2)。一些果皮分泌粘液的物种(如活血丹等), 外果皮细胞分为粘液细胞和非粘液细胞, 粘液细胞较大, 呈长椭圆形或倒卵形, 非粘液细胞较小(图2F)。
中果皮:中果皮为薄壁细胞, 有的细胞壁增厚(如图们黄芩, 图2N)。细胞常1~6层, 或多于6层(如黄芩属, 图2N)。香薷属的少数物种无明显的中果皮细胞(如香薷, 图2D)。中果皮细胞形状常分3种:扁平(如藿香, 图2A); 外侧细胞扁平, 最内层细胞圆形至椭圆形或长方形(如活血丹, 图2F); 细胞近等径(如夏至草属、益母草属等)(图2G)。夏枯草属的中果皮分为两部分, 外层为木化的厚壁细胞, 内侧为近圆形的薄壁细胞(图2L)。大多数物种的维管束较小, 仅由几个导管组成, 通常在横切面不易观察。少数物种维管束呈环状分布于中果皮(如紫苏, 图2I1)或在脊处具一个较大的由导管和纤维构成的维管束(如块根糙苏, 图2J1)。
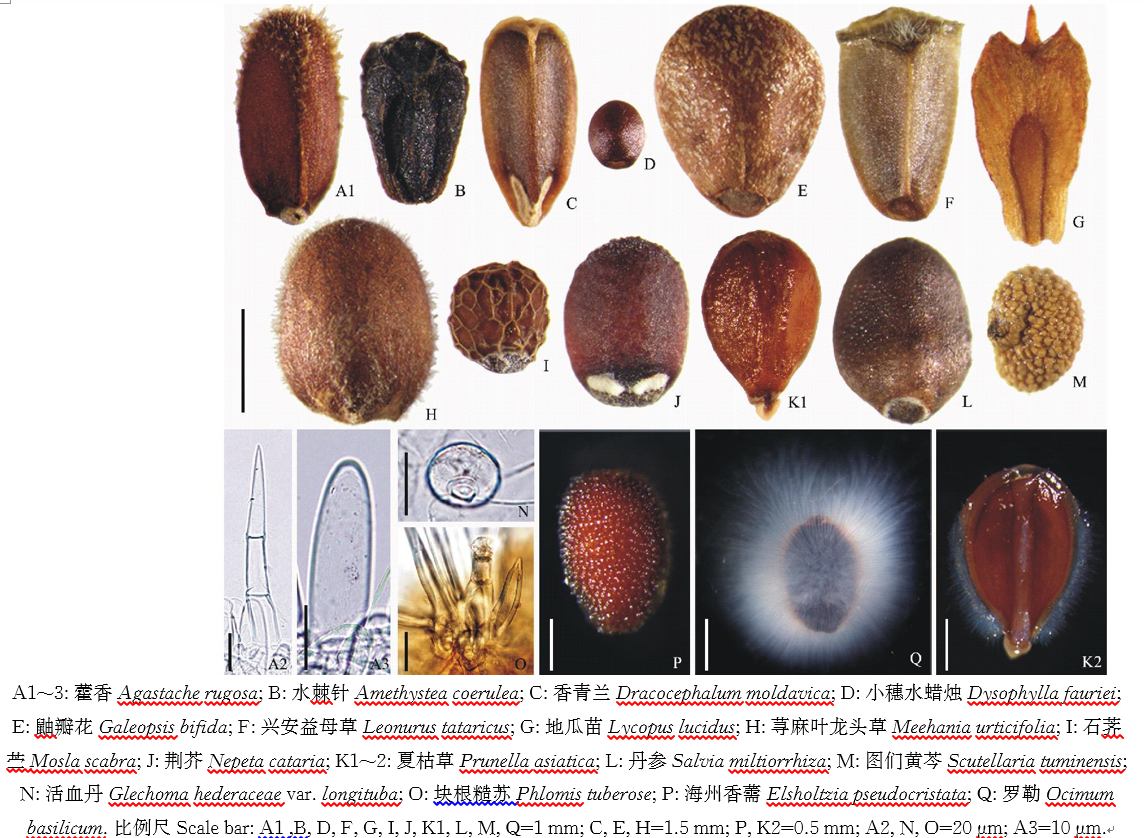 | 图1 东北唇形科果实及毛状体形态Fig.1 Fruit and trichome shapes of Lamiaceae in northeasten China |
厚壁区域:厚壁区域位于中果皮与内果皮之间, 约占果皮厚度的一半(如青兰属和荠苎属等), 多数由一层木化(可见纹孔)的细胞组成(图2B2, P), 少数物种厚壁区域细胞多于一层(如水棘针, 图2B1)。厚壁区域包括石细胞及纤维, 前者具5种类型:1. 两端具分枝, 顶端的分枝长于基部, 中央或近中央处有腔室(图2O), 青兰属和荆芥属等顶端分枝细长呈树枝状, 横切面呈条纹状(图2C1~2); 2. 顶端平, 基部分枝(图2E1, P); 3. 两端近等宽, 切向壁呈脊状突起, 栅栏状排列(图2Q); 4. 星状分枝(图2B2, H2); 5. 近方形, 细胞中央凹陷(图2I1~2)。后者为1或2层, 梭形(块根糙苏, 图2J1~2)。香薷属3种(香薷、海州香薷和木香薷)的厚壁区域的细胞木化(但非石细胞和纤维)并具较大的空腔(图2D)。少数物种(如黄芩属和糙苏属除块根糙苏)无厚壁区域。
内果皮:为一层扁平的薄壁细胞, 偶有2~3层(如内折香茶菜, 图2K)。少数物种内果皮木化, 可见细胞具网状加厚的细胞壁(如紫苏, 图2I1)。细胞近椭圆形、长方形或内切向壁凹陷形成马蹄形。黄芩属的内切向壁着生具极短柄的头状腺毛(图2M, N)。
晶体和丹宁:多数物种具棱柱状的草酸钙单晶, 常分布于厚壁区域细胞的顶端(如鼬瓣花属、益母草属等, 图2E1, G)或中部(图2O, Q), 少数物种存在中果皮(如夏枯草属、黄芩属)。丹宁(经亚铁盐鉴定)分布于果皮的不同区域(如水棘针, 图B1), 某些物种无单宁(如黄芩属)。
 | 图2 东北唇形科果实横切, 示果皮结构、厚壁区域分离的细胞形态Fig.2 The transverse section of fruit of Lamiaceae in northeasten China, showing pericarp structure and separated cell morphology from sclerenchyma region A: 藿香Agastache rugosa; B1~2: 水棘针Amethystea coerulea; C1~2: 光萼青兰Dracocephalum argunense; D: 香薷Elsholtzia ciliate; E1~2: 鼬瓣花Galeopsis bifida; F: 活血丹Glechoma hederaceae var. longituba; G: 细叶益母草Leonurus sibiricus; H1~2: 荠苎Mosla dianthera; I1~2: 紫苏Perilla frutescens; J1~2: 块根糙苏Phlomis tuberose; K: 内折香茶菜Plectranthus inflexus; L: 夏枯草Prunella asiatica; M: 黄芩Scutellaria baicalensis; N: 图们黄芩S. tuminensis; O: 风车草Clinopodium chinense var. grandiflora; P: 小穗水蜡烛Dysophylla fauriei; Q: 地瓜苗Lycopus lucidus。c: 晶体Crystal; en: 内果皮Endocarp; ex: 外果皮Exocarp; f: 纤维Fibre; g: 腺毛Glandular trichome; me: 中果皮Mesocarp; mc: 粘液细胞Mucilaginous cells; nc: 非粘液细胞Non-mucilaginous cells; p: 纹孔Pit; sc: 厚壁区域Sclerenchyma region; vb: 维管束Vascular bundle. 比例尺Scale bar: A, B1, C2, D, H1~2, K, M, J2=10 μ m; C1, E1~2, F, G, I1, J1, N=20 μ m; L=50 μ m; B2, O, P, Q, I2=5 μ m. |
1. 果皮中有草酸钙结晶 2
1. 果皮中无草酸钙结晶 15
2. 无厚壁区域或仅具1~2层纤维 3
2. 有厚壁区域 4
3. 果实为椭圆形或卵圆形, 表面密被突起 黄芩属 Scutellaria
3. 果实为三棱形, 表面光滑或仅顶端密被腺毛和非腺毛 糙苏属 Phlomis
4. 中果皮包括厚壁细胞和1层薄壁细胞, 晶体位于中果皮的薄壁细胞 夏枯草属 Prunella
4. 中果皮不具厚壁细胞, 晶体位于厚壁区域 5
5. 晶体位于厚壁区域细胞中部 6
5. 晶体位于厚壁区域细胞顶端邻近中果皮 8
6. 中果皮细胞1层 风轮菜属 Clinopodium
6. 中果皮细胞多于1层 7
7. 果实顶端圆滑 薄荷属 Mentha
7. 果实顶端成锐齿状, 腹面具腺毛 地瓜苗属 Lycopu
8. 果实三棱形, 2个斜平面光滑 9
8. 果实三棱形, 2个斜平面凹陷, 有一面具裂隙 水棘针属 Amethystea
9. 果实顶端平截 10
9. 果实顶端圆形 11
10. 着生面为圆形 野芝麻属 Lamium
10. 着生面为三角形 益母草属 Leonurus
11. 外果皮浸水后产生粘液 罗勒属 Ocimum
11. 外果皮浸水后不产生粘液 12
12. 果实腹面突起形成锐脊 夏至草属 Lagopsis
12. 果实腹面突起形成的脊平钝 13
13. 果实近圆球状三棱形 水蜡烛属 Dysophylla
13. 果实倒卵状三棱形 14
14. 果实表面光滑 水苏属 Stachys
14. 果实表面具大理石的花纹 鼬瓣花属 Galeopsis
15. 果实为圆球形 16
15. 果实不为圆球形 18
16. 表面具网状纹饰 17
16. 表面光滑, 无网状纹饰 百里香属 Thymus
17. 腹面不具毛状体, 中果皮细胞3~6层, 近等径 紫苏属 Phlomis
17. 腹面具毛状体, 中果皮细胞1~2层, 扁平 荠苎属 Mosla
18. 果实表面具毛状体 19
18. 果实表面不具毛状体 22
19. 内果皮细胞1层 20
19. 内果皮细胞1~4层 香茶菜属Plectranthus
20. 毛状体为腺毛和非腺毛 龙头草属 Meehania
20. 毛状体为腺毛或非腺毛 21
21. 毛状体为腺毛, 中果皮最内层细胞近圆形 活血丹属 Glechoma
21. 毛状体为非腺毛, 中果皮最内层细胞长方形 藿香属 Agastache
22. 着生面为直线形或“ V” 形 23
22. 着生面不为直线形或“ V” 形 24
23. 中果皮细胞多于2层 青兰属 Dracocephalum
23. 中果皮为1~2层 荆芥属 Nepeta
24. 果实表面光滑 裂叶荆芥属 Schizonepeta
24. 果实表面具突起 25
25. 中果皮外侧细胞扁平, 最内层细胞较大, 近长方形 鼠尾草属 Salvia
25. 中果皮外侧细胞扁平, 最内层细胞为椭圆形; 或无明显的中果皮 香薷属 Elsholtzia
Ryding[18]认为地瓜苗属为单系类群, 我们的研究显示地瓜苗属的果实具马蹄形衣领状的翅, 顶端具锐齿(图1G)等独特特征明显不同于该科的其他物种。位于穗花组宽苞亚组的密花香薷和细穗香薷有中果皮和两端具分枝的石细胞, 位于穗花组狭苞亚组的木香薷与位于香薷组的香薷和海州香薷则无明显的中果皮, 厚壁区域细胞木化具较大的空腔, 因而作者认为厚壁区域的特征可作为分类的依据。Jang等[33]观察到密花香薷花粉粒的特有结构(具穿孔的覆盖层和不分支的柱状层), 支持将宽苞亚组独立为组。Budantsev和Lobova[21]将龙头草属和活血丹属放在一个类群, 研究中首次观察到这两个属果实表面存在极短柄的头状腺毛(图1N), 而这种腺毛在荆芥族的其他类群没有观察到。紫苏属与荠苎属的果实表面具网状纹饰, 着生面为圆形, 厚壁区域中无晶体等相似的结构特征支持吴征镒[1]将二者置于紫苏亚族中, 二者较近的亲缘关系也曾得到花粉、果实表面纹饰、雌雄配子体发育及胚胎发生研究的支持[34, 35]。吴征镒[1]依据花萼、花瓣及雄蕊的形态特征将夏枯草属置于夏枯草亚族, 然而, 本研究发现, 夏枯草属中果皮外侧具木化加厚的细胞等特征与其他亚族的类群有很大区别, 夏枯草亚族的系统学有待于进一步深入研究。分子系统学显示荔枝草组应移出荔枝草亚属, 作为独立的亚属, 并认为丹参组应置于鼠尾草亚属[36]。然而, 王开发和黄秀兰[37], 杨德奎等[38]认为荔枝草与丹参花粉的形态及表面纹饰相似, 本研究也显示二者具相似的果皮结构。因此, 荔枝草组与丹参组的关系需要进一步研究。图们黄芩近并头黄芩或为并头黄芩与沙滩黄芩的中间类型[1], 研究显示图们黄芩果实突起的缝隙间具丰富的腺毛, 而并头黄芩与沙滩黄芩无腺毛。黄芩属果实表面密被突起, 中果皮细胞壁轻微木化加厚, 并含有晶体, 无厚壁区域和内果皮具极短柄的头状腺毛等特征不同于唇形科的其他类群, 故支持吴征镒[1]将其放在黄芩亚科。唇形科厚壁区域的晶体被认为是保守的特征, 具有分类学价值[17, 18]。
Moon等[12]指出粘液质常可作为唇形科一些属, 如Dicerandra, Hedeoma, 夏枯草属(Prunella), Thymbra和新塔花属(Ziziphora)的识别特征。粘液的产生与物种生境相关, 干燥地区的物种比湿润环境中的物种更常产生粘液[20], 这一点在本研究中得到证实。果实表面具有网状纹饰或被毛的物种(如荠苎属和龙头草属)不产生粘液, 与Ryding[20]的报道一致。Bü yü kkartal等[23]认为粘液可以将果实固着在土壤中, 其进化与生境相适应。Ryding[20]推测在干旱环境下, 粘液可以储存或吸收水分。
本研究深入揭示了东北唇形科果实的外形、毛状体的类型、着生面的形状和果皮的结构, 并显示这些特征可作为属的分类依据, 完善了东北唇形科果实形态学的研究, 为进一步深入研究中国唇形科形态、分类及系统学提供了形态学基础, 从而也为分子系统学对这一类群的研究提供了基础资料。
致谢:作者感谢中国科学院沈阳应用生态研究所标本馆负责人对本研究提供的支持及帮助。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [19] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [35] |
|
| [37] |
|
| [38] |
|