


{kind=link}
{kind=link}
{kind=link}
{kind=link}
甘蓝型与白菜型冬油菜杂交F1代植物学性状及自交亲和性分析
[赵艳宁1  , 米超
, 米超1 , 孙万仓1, * , 刘自刚1 , 武军艳1 , 方彦1 , 李学才1 , 刘海卿1 , 董云2 ]
, 米超, 刘自刚|
|


作者简介:赵艳宁(1988-),女,甘肃会宁人,硕士.E-mail:zjnn228@163.com
通过甘蓝型冬油菜与北方白菜型冬油菜及其正反交杂种F1植物学形态,生长习性,自交亲和性的比较,分析杂种抗寒性,植物学形态,生长习性,自交亲和性的变化.选择白菜型冬油菜陇油9号与甘蓝型冬油菜Vision组配的(陇油9号×Vision),(Vision×陇油9号)甘白杂交正反交组合,通过亲本与杂种形态比较,越冬率统计,套袋自交以及花粉-柱头互作等,对杂种抗寒性,植物学形态,生长习性,自交亲和性作出评价.陇油9号与Vision杂交,杂种F1植物学形态,干物质积累特性为中间型,生长习性和生长点下凹程度均偏白菜型.F1正,反交越冬率分别为68.75%和65.00%,根长为14.83和14.30 cm,根颈直径为6.08 和5.67 mm.自交亲和性为:Vision>F1 (Vision×陇油9号)>F1 (陇油9号×Vision)>陇油9号,其相应的自交亲和指数为20.83,1.09,1.06,0.87;杂种及其亲本的自交亲和性的大小与其花粉在柱头上的萌发数量与萌发速度相一致.陇油9号与Vision杂交,杂种F1抗寒性强于甘蓝型冬油菜,但较白菜型冬油菜弱.同时,白菜型冬油菜与甘蓝型冬油菜杂交,杂种F1代自交亲和,但杂种的自交亲和程度远远低于甘蓝型冬油菜,而且F1杂种的自交亲和性因杂交组合方式而有差异.
, MI Chao, LIU Zi-GangChanges in cold tolerance, botanical morphology (BM), growth habit (GH) and self-compatibility (SC) of Brassica napus, Brassica rapa and their hybrids were compared. Vision ( B. napus), Longyou 9 ( B. rapa) and their reciprocal crosses progenies were used in the study. The cold tolerance, BM, GH and SC of the hybrids were evaluated by comparing BM, winter performance, bagging selfing and pollen-stigma interactions with that of the parent lines. The BM and dry matter accumulation of F1 hybrids (Vision×Longyou 9) was intermediate while GH was similar to B. rapa. Winter survival of F1 plants in reciprocal crosses were 68.75% and 65.00%. Root length of reciprocal crosses in F1 plants were 14.83 and 14.30 cm. Crown diameter of F1 hybrids from reciprocal crosses were 6.08 and 5.67 mm. The SC was highest in Vision and declined in order; F1 (Vision×Longyou 9), F1 (Longyou 9×Vision), Longyou 9 with SC indices being 20.83, 1.09, 1.06 and 0.87, respectively. The self-compatibility of parents and F1 plants was consistent with the level of pollen germination. The cold tolerance of hybrids was stronger than B. napus but weaker than B. rapa. The F1 hybrids were self-compatible, but the degree of the SC was much less than that of B. napus.
超强抗寒白菜型冬油菜(Brassica rapa)品种的选育成功, 解决了冬油菜抗寒性问题, 使冬油菜安全越冬有了保障[1].但白菜型冬油菜存在产量低, 茎秆强度与株型差, 易倒伏以及品质不良等问题, 因此, 强抗寒白菜型冬油菜品种的品质及农艺性状改良成为育种工作者面临的重要课题.甘蓝型冬油菜(Brassica napus)则具有产量高, 农艺性状优良, 品质优良等特性, 但抗寒性差.通过甘蓝型冬油菜与北方白菜型冬油菜杂交将甘蓝型冬油菜的优良性状导入白菜型冬油菜或将白菜型冬油菜的优良抗寒性导入甘蓝型冬油菜, 选育品质优良, 农艺性状好的白菜型冬油菜或抗寒性优良的甘蓝型冬油菜, 对北方冬油菜生产具有重要意义.甘蓝型冬油菜与北方白菜型冬油菜杂交存在不亲和问题, 杂交后代多偏向甘蓝型冬油菜, 且抗寒性会严重下降.因此, 研究甘蓝型与白菜型冬油菜杂交后代的有关性状和抗寒性, 亲和性的变异是开展甘蓝型冬油菜与白菜型冬油菜的远缘杂交育种的关键环节.远缘杂交是培育油菜新品种的重要途径.刘汉东[2]在远缘杂交后代的变异与选择中发现, 甘蓝型油菜与远缘亲本杂交的亲和性, 表现为亲和关系由近及远, 杂交当代的结角率随之下降, 且以甘× 白组合后代有利变异较多.刘忠松[3]研究发现芥菜型油菜做母本, 甘蓝型油菜和芥菜型油菜种间杂交种可交配性强, 容易获得杂交种子和真杂种.F1代种子发芽率, 成苗率及花粉育性均相对较高, 平均株高低于双亲; 而甘芥杂交较难获得杂种, 且F1代自交亲和性差, 结实率低, 平均株高介于双亲之间[4], 且在芥甘杂种后代中研究发现来自甘蓝型亲本的遗传物质比来自芥菜型亲本的遗传物质在自交后代中丢失更快[5].牛妍等[6]以芥菜型油菜作母本, 白菜型油菜作父本的杂交组合较易获得杂交种子, 杂种F1植株营养生长具有较明显的杂种优势, 但花粉完全不育, 以白菜型油菜回交获得的BC1 F1植株间表型差异明显.付绍红等[7]对甘白油菜种间杂交后代进行加倍处理后发现, 植株在染色体数目, 植株形态, 花器官, 生殖器官上都明显区别于非加倍杂交植株.可以在一定程度上提高杂交后代自交结实, 获得正常的 F2代植株.Liu和Liu[8]研究发现, 油菜远缘杂交普遍存在杂交不亲和现象, 表现为远缘杂交时花粉难以附着在柱头上; 花粉粒不能正常萌发, 花粉管不能正常伸长.近年来胼胝质荧光反应已经广泛用于花粉和雌蕊相互作用的研究[9, 10, 11, 12, 13, 14, 15], 但对甘蓝型冬油菜与北方白菜型冬油菜远缘杂交方面的研究尚未见报道.本研究以甘蓝型冬油菜Vision和强冬性白菜型冬油菜陇油9号及远缘杂交F1代为材料, 对白菜型冬油菜与甘蓝型油菜远缘杂种F1代的亲和性, 抗寒性, 苗期形态特征进行了研究.旨在探明甘蓝型冬油菜和白菜型冬油菜种间杂种F1代主要性状与自交亲和性变化, 为选育具有抗寒性强, 产量高, 农艺性状优良的适宜北方寒旱区的冬油菜新品种提供依据.
参试材料为白菜型冬油菜陇油9号(Longyou 9), 甘蓝型冬油菜Vision, 杂交后代材料F1 (陇油9号× Vision), F1 (Vision× 陇油9号), 种子均由甘肃农业大学农学院提供.
试验于2013-2014年在甘肃农业大学农学院黄家滩试验基地进行, 材料于2013年8月25日播种, 开沟条播, 行距20 cm 株距10 cm, 实验材料采用常规管理.
1.2.1 形态特征和生物学特性调查 2013年10月下旬, 对陇油9号, Vision, F1(陇油9号× Vision)和F1 (Vision× 陇油9号)群体相关形态特征和生物学特性进行记载[16].
1.2.2 干物质积累的测定 2013年11月上旬, 分别从陇油9号, Vision, F1(陇油9号× Vision), F1 (Vision× 陇油9号)中随机挑选5个单株, 挖根后整株带回实验室.清水冲洗干净后从生长点处切开, 测定地上部, 地下部鲜重, 分别测定根长和侧根数后并用数显游标卡尺测定不同世代油菜的根颈直径, 然后将不同世代油菜的根和叶80℃ 烘干测定干重.
1.2.3 花粉活性检测 2014 年 4 月20日(早上10:00-11:00)在亲本及F1正反交群体中随机选取5个单株并采集开花当天花朵带回实验室, 将花药放在载玻片上, 用解剖针碾压花药, 释放出花粉粒, 滴一滴清水, 再滴一滴I2-IK染色液, 在显微镜下观察, 统计花粉活力并拍照.
1.2.4 授粉处理及结角率, 结实率的调查 盛花期选择长势健壮, 枝量, 花序量相当的单株5株, 每个单株选取4个花序, 每个花序选留即将开放的花蕾10~15朵, 其余摘除, 然后套袋(试验期间平均气温为 10~25℃ ).每株自花授粉3个花序, 用于角果成熟后统计结实率和种子数; 1个花序用于套袋自交处理.授粉7 d后, 去掉纸袋并调查各处理的结角率; 角果成熟后调查其结实率, 自交亲和指数.自交亲和指数计算公式为:自交亲和指数=自交结实种子总数/套袋自交总花蕾数, 按照自交亲和指数> 1.00者属于自交亲和, 自交亲和指数< 1.00者为自交不亲和的标准统计[17].
1.2.5 花粉管行为的荧光显微观察 在F1 (陇油9号× Vision)和(Vision× 陇油9号), 甘蓝型冬油菜Vision, 白菜型冬油菜陇油9号群体中随机选择3株, 每个单株选取4个花序, 每个花序选留即将开放的花蕾10~12朵, 其余摘除, 自花授粉后套袋处理.分别在授粉 0, 0.5, 1, 1.5, 2, 4, 6, 8, 12, 24 h 后取样(每个样品 8 根花柱), FAA 固定液(5 mL 福尔马林, 5 mL冰乙酸, 90 mL 70%乙醇)固定后带回实验室.将固定后的花柱用清水冲洗后转移至2 mL离心管中, 用 8 mol/L的 NaOH室温软化16 h, 之后再用0.5%的水溶性苯胺蓝溶液染色 4 h.花柱压片参照Williams等[18]的方法, 花柱整体压片.选取基本等长的已染色花柱, 用镊子去掉花瓣并将其从花柱基部取下, 放在载玻片上, 滴 1 滴甘油, 用盖玻片压片, 在Olympus BHF荧光显微镜下观察花粉在柱头上的粘合, 萌发及花粉管的生长情况并照相.
采用Microsoft Office 2007 软件制表, SPSS 19.0软件对花粉活力镜检结果及自交亲和指数进行显著性分析.
2.1.1 甘白杂交后代与亲本主要植物学性状分析 越冬率是评价植物抗寒性最直观的田间测定指标, 白菜型冬油菜陇油9号的越冬率最高, 为80%; 甘蓝型冬油菜Vision的越冬率最低, 为45.61%; F1 正反交越冬率介于亲本间, 分别为68.75%和65.00%, 正反交差异不大(表1).Vision在心叶色, 叶色, 真叶刺毛, 叶面蜡粉等各形态指标方面均与陇油9号差异较大, Vision生长点在地表之上, 直立生长; 陇油9号生长点在地表之下, 匍匐生长.
| 表1 甘白杂交F1代及亲本形态特征表现 Table 1 Morphological traits of parents and F1 hybrids crosses B. napus with B. rapa |
一般而言, 在冬油菜越冬前心叶色越深, 其抗寒性相对较强.心叶的颜色在F1代正反交中均出现了分离且分离单株比为黄:绿:紫=1:4:10, F1代正反交中, 心叶色为紫色的单株增多, 因此F1代无论正反交较甘蓝型亲本Vision其抗寒性均增强, 这与越冬率统计的结果相一致; F1叶片刺毛性状有分离, 而且因正反交有差异, F1 (陇油9号× Vision)叶片刺毛有:无=3:12, 而反交组合 (Vision× 陇油9号)叶片均表现为无刺毛; F1叶片蜡粉正反交也有差异, (陇油9号× Vision)叶片蜡粉有与无单株之比为5:10, 而(Vision× 陇油9号)叶片蜡粉有与无单株之比为14:1; 甘蓝型与白菜型冬油菜远缘杂交F1生长习性也有分离, F1代无论正反交, 生长习性均出现匍匐生长和半直立生长两种习性, 生长习性均偏向亲本陇油9号的趋势, F1代正反交的抗寒性相对于Vision有所提高; 生长点位置, F1(陇油9号× Vision)生长点地表:地上:地下=9:4:2, F1(Vision× 陇油9号)生长点地表:地上=9:6, 没有出现生长点凹的后代.可见, 以陇油9号做母本时生长点凹陷的趋势更强, 这一结果也从越冬率测定结果得到了验证.
2.1.2 花期和角果期主要形态特征 由图1可见, 白菜型冬油菜亲本陇油9号薹茎叶为全抱茎, 叶色较浅, 叶面无蜡粉(A), 开放的花朵位于花蕾的上方(C); 甘蓝型冬油菜亲本Vision的薹茎叶为半抱茎, 叶色较深, 叶面被蜡粉(B), 开放的花朵位于花蕾的下方(D); (陇油9号× Vision)及反交组合(Vision× 陇油9号)的花蕾与花朵的位置有3种类型, 即花朵在上, 中间型, 花蕾在上(E1, E2, E3), 薹茎叶着生状态也出现了3种类型, 即全抱茎型, 中间型, 半抱茎型(F1, F2, F3), (陇油9号× Vision) E1:E2:E3=1:7:3, F1:F2:F3=1:9:1.但在反交组合(Vision× 陇油9号)中, E1:E2:E3=1:5:5, F1:F2:F3=1:6:4, 说明无论白菜型冬油菜还是甘蓝型冬油菜做母本, 杂种后代的花蕾与花朵的相对位置处于中间型, 并呈偏母本型, 薹茎叶形状偏母本, 但叶片颜色均偏甘蓝型亲本.陇油9号角果长度为5.5~6.5 cm, 角粒数为26~32粒; Vision角果长度为4.5~5.5 cm, 角粒数为24~30粒; (陇油9号× Vision)及反交组合(Vision× 陇油9号)的F1则均为萝卜角果(G), 角果长度为3.0~3.5 cm, 角粒数为10~12粒; 甘白杂种或白甘杂种的角果形态, 角果长度, 角粒数等均为中间型, 且结籽较少, 亲和性低于双亲.
 | 图1 甘白杂交各时期主要形态特征A:陇油9号薹茎叶; B:Vision薹茎叶; C:陇油9号花朵着生状态形态; D:Vision花朵着生状态形态; E:F1代花朵着生状态形态; F:F1薹茎叶 G:陇油9号, Vision, F1 (陇油9号× Vision), F1 (Vision× 陇油9号) 角果形态.Fig.1 Morphological characteristics of B. napus and B. rapa and F1 hybrids in main periodsA: Longyou 9 stem-leaf; B:Vision stem-leaf; C:Longyou 9 flower growth pattern; D:Vision flower growth pattern; E:F1 flower growth pattern; F:F1 stem-leaf; G:Longyou 9, Vision, F1 (Longyou 9× Vision), F1 (Vision× Longyou 9) silique characters. |
Vision冬前地上部发育较快, 长势旺盛, 而陇油9号冬前地上部生长缓慢, 长势较弱.由表2中干物质的积累情况可以看出, 陇油9号侧根数较少, 为直根系, Vision的侧根数较多, 为须根系.(陇油9号× Vision)的侧根数介于两亲本之间且偏向于白菜型亲本, 反交组合(Vision× 陇油9号)的侧根数介于两亲本之间且偏向于甘蓝型亲本.Vision地下部鲜重是地上部鲜重的 23.6%, 而陇油9号地下部鲜重是地上部鲜重的62.3%; F1 (陇油9号× Vision)与F1 (Vision× 陇油9号)的地下部鲜重是地上部鲜重30%左右, 说明在越冬前Vision叶片生长较白菜型冬油菜陇油9号快, 而陇油9号较Vision的根系发育快, (陇油9号× Vision)和(Vision× 陇油9号)F1正反交杂种的叶片与根系生长均介于两个亲本间, 且(陇油9号× Vision)生长较快.(陇油9号× Vision)与反交组合(Vision× 陇油9号)的根长, 根颈直径, 根部干物质重量, 根冠比均大于甘蓝型冬油菜小于白菜型冬油菜.结合表1中结果分析发现, Vision冬前地上部发育较快, 长势旺盛, 越冬率较低, 抗寒性差, 而陇油9号冬前地上部生长缓慢, 长势较弱, 越冬率反而高, 抗寒性好, 种间杂种根冠比和越冬率均处于中间值.因此, 越冬率较高的品种在越冬前根系发育较越冬率低的品种快, 干物质积累量较大.可见, 白菜型冬油菜陇油9号抗寒性优于甘蓝型冬油菜Vision也优于甘白杂种(陇油9号× Vision)和(Vision× 陇油9号), 与其苗期的根系生长和干物质积累特点是有密切关系的.
| 表2 甘白杂交F1代及亲本苗期干物质积累 Table 2 Dry matter accumulation of B. napus and B. rapa and F1 hybrids |
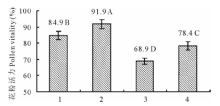
2.3.1 甘白杂交后代花粉活力 I2-IK染色镜检结果(图2)表明, Vision花粉可染率高, 平均可染率达91.9%(图3), 花粉粒着色普遍较深.陇油9号花粉可染率低于Vision, 为84.9%, 花粉粒着色较深, 但存在发育不规则花粉与三核花粉(图2A, C). 甘蓝型与白菜型冬油菜杂交F1, 无论白甘杂种还是甘白杂种, 花粉大小不一致, 也出现了不规则花粉和三核花粉, 且F1 (陇油9号× Vision)花粉平均可染率仅68.9%, 而Vision× 陇油9号花粉平均可染率为78.4%.说明甘蓝型冬油菜与白菜型冬油菜杂交, 杂种花粉活力均低于亲本, 但以甘蓝型冬油菜作母本F1花粉活力高于其反交组合, 这与甘白远缘杂交F1代自交结实率的结果相一致(表3).
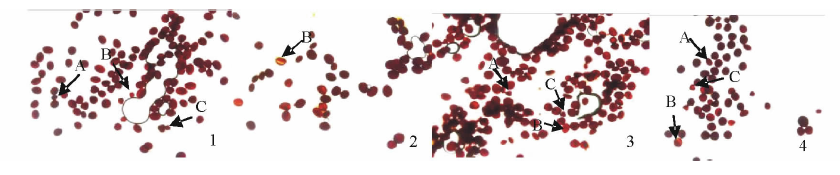 | 图2 亲本及F1代花粉I2-IK染色结果1.陇油9号 Longyou 9; 2.Vision; 3. F1(Longyou 9× Vision); 4. F1(Vision× Longyou 9); 下同.The same below.A:不规则花粉粒Irregular pollen grains; B:染色较浅花粉Lighter staining pollen grains; C:三核花粉Three nuclear pollen grains.Fig.2 Results of pollens staining by I2-IK of parents and F1 hybrids |
 | 图3 甘白杂交花粉活力不同字母表示差异显著(P< 0.05).Fig.3 Pollen vitality of B. napus and B. rapa and F1 hybridsDifferent letters above bars indicate significant differences at P< 0.05. |
2.3.2 甘白杂交后代自交亲和指数 盛花期对亲本及杂种进行套袋处理, 共套袋处理118个单株, 1487朵花.Vision所有单株均自交亲和, 结角率达95%, 平均自交亲和指数达20.83; 陇油9号均表现出自交不亲和, 结角率仅4%, 平均自交亲和指数0.87.杂种F1代无论正反交, 自交亲和株率, 结角率较白菜型冬油菜陇油9号均大幅提高, 结角率达60%左右, 亲和指数分布在1.06~1.09之间, 但远远低于甘蓝型冬油菜亲本; 甘白杂交F1自交亲和性程度也因杂交的组合方式而有较大差异, (陇油9号× Vision)的自交亲和株率(自交亲和株率依据表3中自交亲和指数变异范围的亲和单株数计算得出), 结角率, 自交亲和指数分别为39.3%, 61%, 1.06, (Vision× 陇油9号)自交亲和株率, 结角率, 自交亲和指数分别为60%, 67%, 1.09, 可见母本对杂种自交亲和性的影响较大(表3).
| 表3 甘白杂交亲本及后代自交亲和个体分布显著性比较 Table 3 Distribution of self-compatible plant and the significance analysis in B. napus and B. rapa and F1 hybrids |
2.3.3 甘白杂交后代自花授粉柱头上花粉萌发及花粉管生长 Vision在自交授粉0 h, 即有少量花粉粘附在柱头表面(图4A-1); 0.5 h后大量花粉粒粘附到柱头表面(图4A-2); 授粉后1 h, 花粉粒体积增大(图4A-3), 极少数花粉粒萌发孔处出现绿色荧光(图4A-4); 1.5 h后约30%的花粉粒萌发(图4A-5); 2 h后大量花粉开始萌发, 长出花粉管(图4A-6); 4~8 h花粉管开始伸长, (图4A-7, 8, 9); 24 h后花粉管长度超过100 μ m(图4A-10).
 | 图4 亲本及F1正反交自花花粉萌发及花粉管生长情况A~D: Vision, 陇油9号, F1 (陇油9号× Vision)及F1 (Vision× 陇油9号)自花花粉萌发及花粉管生长情况; A-1: 授粉后 0 h, 少量花粉粘附在柱头上, 50× ; A-2: 授粉后0.5 h, 大量花粉粘附在柱头表面, 50× ; A-3: 授粉后1 h, 花粉粒体积增大, 200× ; A-4: 授粉后 1 h, 花粉粒萌发孔处出现绿色荧光, 400× ; A-5: 授粉后1.5 h, 部分花粉粒萌发, 200× ; A-6: 授粉后2 h, 大量花粉开始萌发, 200× ; A-7和 A-8: 授粉后4, 6 h, 花粉管开始伸长, 200× ; A-9: 授粉后8 h, 花粉管开始伸长, 400× ; A-10: 授粉后 24 h, 花粉管继续伸长, 长度超过100 μ m, 400× ; B-1: 授粉后 0 h, 柱头上无花粉粒, 50× ; B-2: 授粉后0.5 h, 少量花粉粘附在柱头表面, 50× ; B-3: 授粉后1 h, 大量花粉粘附在柱头表面, 100× ; B-4: 授粉后1.5 h, 部分花粉粒萌发孔处出现绿色荧光, 100× ; B-5: 授粉后2 h, 50%花粉粒长出花粉管, 大量胼胝质沉积, 100× ; B-6: 授粉后4 h, 胼胝质反应加剧, 200× ; B-7: 授粉后6 h, 花粉管开始伸长, 胼胝质反应剧烈, 200× ; B-8: 授粉 8 h 后, 花粉管长达50 μ m, 400× ; B-9: 授粉后 24 h, 少部分花粉管尾部缠绕回折, 400× ; C-1: 授粉后 0 h, 柱头上无花粉粒, 50× ; C-2: 授粉后0.5 h, 极少数花粉粘附在柱头表面, 50× ; C-3: 授粉后1 h, 较多花粉粘附在柱头表面, 100× ; C-4: 授粉后1.5 h, 大量花粉粘附在柱头表面, 100× ; C-5: 授粉后2 h, 长出花粉管, 大量胼胝质沉积, 100× ; C-6: 授粉后4 h, 约80%的花粉粒体积变大, 200× ; C-7: 授粉后6 h, 少数花粉粒长出花粉管, 200× ; C-8: 授粉 8 h 后, 大部分花粉粒长出花粉管, 400× ; C-9: 授粉后 24 h, 花粉管伸长, 长度仅30 μ m, 400× ; D-1: 授粉后 0 h, 柱头上粘附较少花粉粒, 50× ; D-2: 授粉后0.5 h, 较多花粉粘附在柱头表面, 50× ; D-3: 授粉后1 h, 少量胼胝质沉积, 100× ; D-4: 授粉后1.5 h, 大量胼胝质沉积, 100× ; D-5: 授粉后2 h, 大部分花粉粒体积增大, 100× ; D-6: 授粉后4 h, 进入萌发高峰期, 大量花粉开始萌发, 200× ; D-7: 授粉后6 h, 花粉管开始伸长, 200× ; D-8: 授粉后8 h, 花粉管长达50 μ m, 400× ; D-9:授粉后 24 h, 少部分花粉管尾部缠绕回折, 400× ; Pg:花粉粒; PT:花粉管; Cr:胼胝质反应; Cp:胼胝质塞.Fig.4 Pollens germination and growth in parents and F1 reciprocal cross self-crossA-D: Pollens germination and growth in Vision, Longyou 9, F1 (Longyou 9× Vision) and F1 (Vision× Longyou 9) self-cross. A-1: 0 h after the cross, a few pollen adheres to the stigma, 50× ; A-2: 0.5 h after the cross, lots of pollen adheres to the stigma, 50× ; A-3: 1 h after the cross, pollen volume increases, 200× ; A-4: 1 h after the cross, green fluorescent appeared at pollen grain hole, 400× ; A-5: 1.5 h after the cross, part of pollen began to germinate, 200× ; A-6: 2 h after the cross, lots of pollen began to germinate, 200× ; A-7 and A-8: 4 and 6 h after the cross pollen tube began to elongate, 200× ; A-9: 8 h after the cross pollen tube began to elongate, 400× ; A-10: 24 h after the cross pollen tube continues to elongate longer than 100 μ m, 400× ; B-1: 0 h after the cross, no pollen adheres to the stigma, 50× ; B-2: 0.5 h after the cross, a few pollen adheres to the stigma, 50× ; B-3: 1 h after the cross, lots of pollen adheres to the stigma, 100× ; B-4: 1.5 h after the cross, most of the pollen grains germinate hole appeared green fluorescence, 100× ; B-5: 2 h after the cross, 50% of pollen began to germinate and lots of calloses emerged around papilla cells, 100× ; B-6: 4 h after the cross, callose reaction increased, 200× ; B-7: 6 h after the cross, pollen tube began to elongate and callose violent reaction, 200× ; B-8: 8 h after the cross, pollen tube longer than 50 μ m, 400× ; B-9: 24 h after the cross, a few pollen tube spiraled, 400× ; C-1: 0 h after the cross, no pollen adheres to the stigma, 50× ; C-2: 0.5 h after the cross a few pollen adheres to the stigma, 50× ; C-3: 1 h after the cross, more pollen adheres to the stigma, 100× ; C-4: 1.5 h after the cross, lots of pollen adheres to the stigma, 100× ; C-5: 2 h after the cross, pollen began to germinate and lots of calloses emerged around papilla cells, 100× ; C-6: 4 h after the cross, more than 80% pollen volume increases, 200× ; C-7: 6 h after the cross, a few pollen grow pollen tube, 200× ; C-8: 8 h after the cross, lots of pollen began to grow pollen tube, 400× ; C-9: 24 h after the cross, pollen tube continues to elongate longer than 30 μ m, 400× ; D-1: 0 h after the cross, a few pollen adheres to the stigma, 50× ; D-2: 0.5 h after the cross, more pollen adheres to the stigma, 50× ; D-3: 1 h after the cross, a few calloses emerged around papilla cells, 100× ; D-4: 1.5 h after the cross, lots of calloses emerged around papilla cells, 100× ; D-5: 2 h after the cross, most of pollen volume increases, 100× ; D-6: 4 h after the cross, enter the germination peak period and lots of pollen began to germinate, 200× ; D-7: 6 h after the cross, pollen tube began to elongate, 200× ; D-8: 8 h after the cross, pollen tube longer than 50 μ m, 400× ; D-9: 24 h after the cross, a small part of the pollen tube tail is spiraled and back to the fold, 400× ; Pg: Pollen grain; PT: Pollen tube; Cr: Callose reaction; Cp: Callose plug. |
陇油9号自花授粉0 h, 柱头上无花粉粒(图4B-1); 授粉0.5 h, 少量花粉粒粘附在柱头上(图4B-2), 授粉后1 h, 大量花粉粒粘附在柱头上, 花粉粒体积增大(图4B-3); 1.5 h后约60%的花粉粒萌发孔处变亮, 即将进入萌发阶段(图4B-4); 2 h后约50%的花粉粒长出花粉管, 柱头乳突细胞中出现大量的胼胝质沉积(图4B-5); 授粉后4 h胼胝质反应加剧(图4B-6); 授粉6 h后, 花粉管开始伸长, 部分花粉管由于受胼胝质阻碍而弯曲生长(图4B-7).授粉8 h后, 花粉管长度达50 μ m(图4B-8); 24 h后, 极少数花粉粒还未萌发, 少部分花粉管尾部出现缠绕, 回折现象; 部分花粉管中长出胼胝质塞(图4B-9).
比较Vision和陇油9号花粉粒在柱头上的萌发动态发现:花粉粒在Vision柱头上发生水合作用的时间大概在1 h, 授粉2 h后进入萌发高峰期, 长出花粉管; 直至观察结束(24 h), 整个花粉萌发, 花粉管生长阶段中胼胝质反应较弱, 花粉管在柱头上能正常生长.相较于Vision, 花粉在陇油9号柱头上粘附时间略滞后, 大概在1.5 h左右; 授粉2 h后柱头乳突细胞中胼胝质大量沉积; 授粉4 h后长出花粉管; 整个观察阶段中都伴有较强的胼胝质反应; 24 h后, 少部分花粉管尾部出现缠绕, 回折现象; 从花粉粒萌发过程中的胼胝质反应程度和花粉管生长过程中的回折现象以及花粉管中出现的胼胝质塞足以说明对陇油9号的授粉受精过程造成了较大影响.
(陇油9号× Vision)杂种, 授粉0 h时柱头上无花粉粒附着(图4C-1); 授粉0.5 h时有极少数花粉粒粘附在柱头上(图4C-2), 授粉后1 h开始有较多花粉粒粘附在柱头上, 柱头乳突细胞中出现少量胼胝质, 少数花粉粒体积增大(图4C-3); 1.5 h后, 花粉粒大量粘合在柱头上, 胼胝质反应增强(图4C-4); 2 h后柱头乳突细胞中的胼胝质沉积继续增加, 少量花粉粒萌发孔处变亮(图4C-5); 授粉后4 h, 约80%的花粉粒体积变大, 胼胝质反应最剧烈(图4C-6); 授粉6 h后, 少数花粉粒长出花粉管(图4C-7).授粉8 h后, 大部分花粉粒萌发(图4C-8); 24 h后, 花粉管伸长, 但长度仅30 μ m左右 (图4C-9).反交杂种F1 (Vision× 陇油9号)自花授粉0 h, 柱头上粘合的花粉粒仍然较少(图4D-1); 授粉0.5 h较多的花粉粒粘附在柱头上(图4D-2); 授粉后1 h柱头乳突细胞中出现少量胼胝质, 花粉粒体积增大(图4D-3); 1.5 h后柱头乳突细胞中出现大量胼胝质沉积, 极少数花粉粒萌发长出花粉管(图4D-4); 2 h后花粉粒大量粘合在柱头上, 大部分花粉粒体积增大(图4D-5); 授粉后4 h, 胼胝质反应最剧烈, 绝大部分花粉粒体积膨胀, 萌发孔处变亮, 进入萌发阶段(图4D-6); 授粉6 h后, 一半以上花粉粒长出花粉管, 花粉管开始伸长(图4D-7).授粉8 h后, 花粉管开始伸长(图4D-8); 24 h后, 花粉管伸长, 长度达50 μ m以上, 部分花粉管中长出胼胝质塞, 少数花粉管出现缠绕现象(图4D-9).可见, 甘蓝型与白菜型冬油菜杂交, 以甘蓝型冬油菜作母本, 在相应时段内, 杂种柱头上花粉附着速度, 附着量, 花粉萌发时间, 萌发量要大于白菜型冬油菜作母本的处理.
白菜型冬油菜与甘蓝型冬油菜种间杂种F1有分离, 生长习性偏白菜型冬油菜, 大多数种间杂种匍匐生长, 以白菜型油菜作母本生长点凹陷比甘蓝型油菜作母本更明显, 生长点凹陷是白菜型冬油菜之所以抗寒的植物学形态保障; 叶色及叶面蜡粉特性偏向于白菜型冬油菜, 而叶片刺毛性状则偏向于甘蓝型亲本; 杂种后代无论正反交其花蕾与花朵的相对位置以及叶形均呈现出偏母本性状, 薹茎叶半抱茎, 颜色均偏向于甘蓝型亲本; 杂种角果短小, 呈萝卜角果, 果喙较短, 角果长度及每角粒数等为中间型; 花期也介于两亲本间, 较白菜型亲本晚, 较甘蓝型亲本早, 但在整体形态上杂种植株偏甘蓝型亲本.杂种植株营养生长茂盛, 叶片较厚, 分枝较多, 表现出较明显的杂种优势, 根系的形态也介于甘蓝型与白菜型亲本之间.甘白杂种的抗寒性介于亲本间, 抗寒性的变化可能与形态特征的变化有关, 这与前人对冬油菜幼苗习性的研究结果相一致[19].
根冠比是判断植物干物质积累的一项重要指标, 而越冬率是判断植物抗寒性的特征指标.研究表明, 甘蓝型冬油菜与白菜型冬油菜冬前生长和干物质积累特点是完全不同的, 甘蓝型冬油菜冬前生长十分迅速, 主要是长叶, 而白菜型冬油菜则生长缓慢, 主要是长根.越冬率调查结果显示, 越冬率高低也是Vision< F1代< 陇油9号, 这与结缕草属植物抗寒性研究结果一致[20], 没有表现出母性遗传的特征.并且, 甘白远缘杂交F1代生长速度, 干物质的积累动态介于两亲本间, 无论是白菜型油菜做母本还是甘蓝型油菜做母本, 根冠比规律均为:Vision< F1代< 陇油9号, 说明陇油9号及F1正反交后代在越冬前根系较Vision发育快.说明(白菜型× 甘蓝型)和(甘蓝型× 白菜型)杂交F1代抗寒性较甘蓝型亲本有所提高, 与其干物质积累特点有关.
本研究表明, 甘蓝型冬油菜与白菜型冬油菜杂交, 杂种F1是自交亲和的, 这种亲和性无论在群体亲和株率, 结角率, 亲和指数, 花粉在柱头上的萌发情况均得到体现, 但杂种的自交亲和程度远远低于甘蓝型冬油菜.甘白杂种群体亲和株率, 结角率, 亲和指数小于甘蓝型冬油菜, 同时, 甘白杂种的自交亲和性因杂交组合方式而有差异, (Vision× 陇油9号)自交亲和性> (陇油9号× Vision), 亲本及杂种F1的自交亲和指数大小为:Vision> F1 (Vision× 陇油9号)> F1 (陇油9号× Vision) > 陇油9号, 其相应的自交亲和指数为20.83, 1.09, 1.06, 0.87.杂种及其亲本的自交亲和性的大小与其花粉在柱头上的萌发数量与萌发速度相一致.前人研究表明, 有性生殖中植物花的性表达特征与植物适合度间有着内在的联系[21], 虽然很多植物在种间杂交时果实能膨大坐果, 但不一定有可育种子[22].本试验中, 甘白种间杂交后代自交结实率仅为1%左右, 而且每角粒数很少, 平均仅为0.6~1.3 粒, 所以用结实率和每角粒数两个指标能更准确反映甘白F1代正反交的亲和性.王文和等[23]认为柱头乳突细胞是阻碍花粉管生长的第一道屏障, 花粉落到柱头后, 需经历萌发, 花粉管在花柱内的生长及穿过珠孔等过程, 才能实现受精[24, 25].而本试验中陇油9号自花授粉4 h, 柱头乳突细胞中出现大量胼胝质沉积, Vision则在授粉后0.5 h出现少量胼胝质; F1 (陇油9号× Vision)和F1 (Vision× 陇油9号)分别在自花授粉4和2 h后出现大量胼胝质.巩振辉等[26], 王幼平等[27], 孙万仓等[28]研究发现十字花科植物属于受精后不亲和类型, 孟金陵等[29]指出花粉粒在柱头表面粘合后在乳突细胞内产生胼胝质, 花粉管受阻于乳突细胞, 这是十字花科植物中由雌蕊S等位基因编码的S位点糖蛋白(SLG)和S受体激酶决定的, 它们可能与磷酸化和去磷酸化参与了的某种信号传递有关, 最后导致自交花粉的抑制[30].并且远缘杂交后代亲和性的高低与母本自交亲和性的高低有密切关系.而本试验中, 陇油9号自花授粉4~6 h, 萌发量最大, Vision自花授粉2 h后萌发量达到最大, F1 (陇油9号× Vision)自花授粉8 h后萌发量最大, 而F1 (Vision× 陇油9号)自花授粉4 h后花粉萌发量最大.总体来说, 自花授粉后柱头上的花粉均以单粒形式存在[31], 萌发率随授粉时间的延长而增加, 这与陈迪新[32]对梨花粉萌发的结果相一致.本试验观察到, 以陇油9号为母本, F1代自花花粉在柱头上萌发率较低, 萌发速率较慢, 花粉黏附在柱头上的时间延迟, 花粉管生长发育迟缓, 且柱头乳突细胞表面存在大量胼胝质沉积, 胼胝质反应强烈, 个别生长发生严重扭曲, 花粉管出现末端膨大, 中间部分折叠, 花粉管内出现胼胝质塞等现象; 这与李守丽等[33]对百合属的研究结果相类似; 而以Vision做母本, F1代自花花粉在柱头上萌发率较高且萌发速率较快, 柱头乳突细胞中的胼胝质出现时间较晚, 数量也较少.
本试验中观察到4个试验群体的自花花粉在柱头上萌发时花粉粒直径均达到25~30 μ m时, 花粉萌发孔处才变亮, 发出绿色荧光, 可以将这一现象视作观察花粉水合作用结束, 开始萌发的标志.因此, 深入研究甘白杂交正反交后代间的花粉萌发, 花粉管生长特点, 便于确定种子的形成和种子的发育状况, 进而为早期世代中选择出亲和性单株奠定基础.此外, 油菜亲和性的大小也受到日照, 温度等环境因素的影响, 长日照, 高温对受精过程起促进作用, 可能是改变了柱头上一些与温度有关的诱导性酶的启动先后顺序, 需对自交亲和性机制做进一步的研究.
本研究表明, 甘蓝型冬油菜与白菜型冬油菜杂交, 杂种F1植物学形态, 干物质积累特性为中间型, 生长习性偏白菜型冬油菜, 抗寒性强于甘蓝型冬油菜, 但较白菜型冬油菜弱.同时, 甘蓝型冬油菜与白菜型冬油菜杂种F1是自交亲和的, 但杂种的自交亲和程度远远低于甘蓝型冬油菜, 而且甘白杂种的自交亲和性因杂交组合方式而有差异, 自交亲和性表现为:(甘蓝型冬油菜× 白菜型冬油菜)> (白菜型冬油菜× 甘蓝型冬油菜).甘蓝型冬油菜, 白菜型冬油菜亲本及杂种F1的自交亲和指数大小为:甘蓝型冬油菜> F1 (甘蓝型冬油菜× 白菜型冬油菜)> F1 (白菜型冬油菜× 甘蓝型冬油菜)> 白菜型冬油菜, 其相应的自交亲和指数为20.83, 1.09, 1.06, 0.87, 杂种F1及其亲本的自交亲和性的大小与其花粉在柱头上的萌发数量与萌发速度相一致.因此, 通过甘蓝型冬油菜与白菜型冬油菜杂交来改良甘蓝型冬油菜的抗寒性是可能的, 对改良白菜型冬油菜的自交亲和性也有积极意义.
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|