


{kind=link}
{kind=link}
{kind=link}
{kind=link}
暗紫贝母花器官特征及有性繁育系统研究
[陈垣1, 2  , 徐博琼
, 徐博琼1 , 郭凤霞1, * , 白刚1 , 张荩凤1 , 张勇3 ]
, 徐博琼, 白刚|
|


作者简介:陈垣(1963-),男,甘肃通渭人,教授,博士生导师。E-mail: chenyuan@gsau.edu.cn
采用田间试验与显微镜检相结合的方法,对漳县人工栽培的暗紫贝母花器构造和传粉生物学进行研究,旨在揭示其花器官特征和有性繁育系统,为暗紫贝母的育种提供科学依据。结果表明,经鳞茎播种栽培3年的暗紫贝母植株单花顶生,偶见2花,花两性,初现花蕾绿色,花暗紫色,有淡黄色格纹,开花期5-6月,单花寿命6~8 d,花钟形,同被花,花被6,2轮,离生雄蕊6枚,5枚紧围雌蕊,1枚外围,子房上位,柱头3裂,短裂片外露,毛状有分泌物。开花前柱头高于雄蕊,开放后雄蕊高出柱头,开花当天花粉量最大,活力最高并维持3 d时间。花被长1.5 cm的绿蕾柱头已具较强可受性。花药开裂与强柱头可受性同步,但后者持续期更长(6 d以上)。单花花粉数约6000粒、胚珠数在160颗以上,花粉胚珠比( P/O)37.5,杂交指数(OCI)为4。传粉媒介为风媒和虫媒,访花者为蜂类、蝇类和蜘蛛,但数量极少,有性繁育系统应属于部分自交亲和,异交,需要传粉者。
To determine the flower characteristics and sexual breeding system for Fritillaria unibracteata, a combination of field tests and microscopy examination was used to study the flower conformation and pollination biology in order to provide scientific information to assist in plant breeding. This species has a single bisexual flower borne at the stem apex. Occasionally there are two flowers when cultivated for three years from bulbs. The immature bud is green and then turns a darker purple covered with light yellow flecks. The flowering period is from May to June but single flowers bloom only 6-8 days. The bell-shaped flower has six perianth segments in two whorls and six apopetalous stamens within, five of which are tightly circumfused to the pistil and the other independent. The epigynous ovary has three short splits on its hairy chapiter which are secretory. The ovary stands higher than the stamens in the young bud, but becomes shorter in mature, open flowers. The pollen quantity, and vigor is maximal on the first day of flowering and pollen is produced for three days. The pistil stigma in a 1.5 cm long green bud was already receptive. Anther dehiscence coincided with the peak in stigma receptivity but stigma receptivity lasted longer (over 6 days) than pollen production. The flower has over 6000 pollen grains and 160 ovules in single flower, resulting in a pollen to ovule (P/O) ratio of 37.5. The out crossing index (OCI) was 4 according to the flower size and behavior. Pollinators included wind and a few wasps, bees, spiders and other miscellaneous insects. Our results suggest that this species should possess a sexual breeding system with partial self-compatibility, heterogamy, and required pollinators.
暗紫贝母(Fritillaria unibracteata)是药材川贝母的主要基原植物之一, 鳞茎入药, 味苦、甘, 微寒, 归肺、心经, 具有清热散结, 化痰止咳等功效, 用于治疗肺热燥咳、散结消痈等症[1]。其鳞茎中有多种生物碱、皂苷和人体必需微量元素等[2, 3]。暗紫贝母野生于海拔3200~4500 m的草地上, 我国主产于四川、甘肃、青海等地, 分布范围十分有限[4]。
然而, 暗紫贝母生长环境特殊, 气候冷凉, 药材生长周期长(4年), 药材产量低, 因采挖过度, 野生资源已趋于枯竭[4], 市场价格昂贵。自然条件下, 暗紫贝母主要依赖鳞茎无性繁殖。由于种子具有明显的后熟和休眠特性, 种子落地后一般很难满足其萌发条件, 自然有性繁殖率很低, 野生驯化栽培时也多以鳞茎繁殖为主, 但采用鳞茎繁殖, 一方面引起种质资源的退化, 另一方面繁殖系数低且消耗了部分商品鳞茎, 成本高, 采用种子繁殖, 其繁殖系数可提高50倍以上[5, 6, 7]。植物的繁育系统是植物内在遗传特性和外界环境相互作用的结果, 是种群有性生殖的纽带, 对植物的遗传变异和进化过程均有着重要的意义[8]。植物的花被特征、花粉活力、柱头可受性及杂交指数(outcrossing index, OCI)等指标均是反映植物有性生殖能力的重要依据[9, 10, 11, 12, 13, 14, 15, 16]。暗紫贝母作为市场紧缺的名贵中药材, 随着野生资源生态环境扰动加大, 现有的资源已难以满足市场的需求, 驯化栽培是保障药源的重要措施, 利用种子繁殖无疑成为保护野生资源, 提高资源利用率的有效途径。尽管前人对不同海拔的暗紫贝母花朵和果实大小进行了比较[4], 并对其组培快繁技术进行了探索[14], 上述研究对暗紫贝母种子繁育策略的制定提供了一定参考, 甘肃农业大学对甘肃贝母(F. przewalskii)种子灌浆特性和种子发芽特性的研究系统而深入, 种子繁殖技术已趋成熟[5, 6, 7], 但至今对暗紫贝母等川贝母基原植物的有性繁育特性的系统研究尚少见报道, 严重限制了川贝母种子繁育。因此, 在适生地对驯化栽培的野生暗紫贝母花被特征及有性繁育特性开展深入研究具有重要意义, 可为川贝母基原植物种子繁育和品种保纯提供科学依据, 有利于促进野生川贝母资源的可持续发展, 实现中药资源的现代化管理。
试验区位于甘肃省定西市漳县南部山区石川乡菜子川村, 东连岷县蒲麻镇, 南与岷县禾驮乡接壤, 西靠四族乡, 北接草滩乡, 地处东经104° 19'41″, 北纬34° 35'29″, 海拔3100 m的山坡, 无霜期126 d, 年平均降雨量500 mm, 属典型高寒阴湿气候, 气候冷凉, 土层深厚肥沃, 适宜暗紫贝母生长发育。试验地为栽培1年当归后撂荒3年的农田。试验材料为2013年野生暗紫贝母鳞茎驯化种植栽培的种株, 经甘肃农业大学中草药栽培与鉴定系陈垣教授鉴定确认为暗紫贝母, 种鳞茎采自四川红原县, 驯化栽培采用撒播, 覆土厚度3 cm, 播种后覆盖遮阳网, 面积0.267 hm2, 2014年返青后揭去遮阳网。
从2015年返青后5月1日暗紫贝母花蕾显现开始, 田间选取具有代表性的20株单花植株挂牌标记, 从现蕾开始至开花结束, 定位每天观察记录单花结构、花粉散出和花朵萎蔫情况, 采用游标卡尺测定花冠长度(corolla length)和花冠直径(corolla diameter)等生长发育指标。以花被全开放为100%标准, 将花被开度划分为4个级别, 即1)闭合, 开花前, 花被尚未开放; 2)微开, 开花当天, 花被开张度25%以下, 用0 d表示; 3)半开, 花被开张度50%左右; 4)开, 花被全开, 开张度达100%。
花粉活力采用张志良[15]的0.5%TTC(2, 3, 5-氯代三苯基四氮唑)方法测定, 即从不同发育阶段的花朵中用镊子轻轻取下1枚花药, 经处理使花粉完全散出于准备好的载玻片上, 滴加1滴0.5%TTC染色, 充分混匀后盖上盖玻片, 置于有湿润滤纸的培养皿中置37 ℃黑暗静置15 min, 然后在光学显微镜(4× )和LY-SUPER HP CCD万能成像系统下观察, 根据花粉染色情况确定花粉活力, 红色表示花粉活力强, 浅红色表示花粉有活力, 但活力弱, 无色或黄色则说明花粉无活力。统计盖玻片视野内红色、粉红色和无色花粉粒数, 每枚雄蕊观察5个视野, 每朵花重复观察3枚雄蕊, 共观察统计15个视野(n=15), 最后计算花粉活率。
花粉活率(pollen vigor rate, %)=视野内淡红和红色花粉粒总数× 100/视野内花粉粒总数
强活力花粉粒率(strong-pollen rate, %)=视野内红色花粉粒数× 100/视野内花粉粒总数
弱活力花粉粒率(weak-pollen rate, %)=视野内粉色花粉粒数× 100/视野内花粉粒总数
参照Dafni等[10]的方法, 采用联苯胺-过氧化氢法对柱头可受性测定。即将联苯胺-过氧化氢反应液(1%联苯胺:3%过氧化氢:水=4:11:22)滴在凹式载玻片上, 然后用镊子小心取出不同发育期花朵的柱头分别浸入反应液中。在显微镜(10× )下观察有无气泡产生。若柱头周围的反应液呈现蓝色并伴有大量气泡出现, 则说明柱头具有可受性, 反之则说明柱头无可受性。每个处理花朵重复测定3次, 每次从5个角度观察。
将花药尚未开裂的绿色花蕾轻轻剥开, 用镊子小心取出未开裂的雄蕊1枚, 将花药挤碎于载玻片上, 滴入0.5%TTC, 盖上盖玻片黑暗染色15 min, 将盖玻片表面用铅笔划分为6格, 在光学显微镜(10× )下统计每格花粉粒数(pollen), 重复3个花蕾。取心皮在体视显微镜下用解剖针划开, 计数胚珠数(ovule), 重复3次。用平均单花花粉量除以单花胚珠数得到P/O值。按Cruden[9]的标准, 计算花粉量-胚珠比(the pollen-ovule ratio, P/O), P/O为2.7~5.4 时, 其繁育系统为闭花受精; P/O为18.1~39.0时, 繁育系统为专性自交(obligate autogamy); P/O为31.9~396.0 时, 繁育系统为兼性自交(facultative autogamy); P/O为244.7~2588.0时, 繁育系统为兼性异交(facultative outcrossing); P/O为2108.0~195525.0时, 繁育系统为专性异交(obligate outcrossing)。
参照任明迅[16]的方法, 按照Dafni[11]的标准, 根据花朵大小和开花行为对繁育系统综合评判, 即首先根据花朵直径确定杂交指数, 若花朵直径< 0.1 cm记为0, 0.1~0.2 cm记为1, 0.2~0.6 cm记为2, > 0.6 cm记为3; 其次以花药开裂时间与柱头可受性之间的时间间隔为依据, 若雌雄同时成熟或雌蕊先熟记为0, 雄蕊先熟记为1; 最后根据柱头与花药的空间位置, 若柱头与花药在同一高度记为0, 空间分离记为1。以上三者之和为杂交指数(outcrossing index, OCI)值。杂交指数越大, 异交程度越高, 越需要传粉者[11, 16]。具体评判标准详见表1。
| 表1 植物繁育系统的评判标准 Table 1 Criterion of plant reproduction system |
采用Excel 2003软件制图, SPSS 11.5软件进行方差分析, 采用Duncan法进行多重比较。图中数据点为平均数± 标准误(mean± SE)。
田间观察表明, 在定西市漳县石川乡驯化栽培的野生暗紫贝母为百合科贝母属多年生草本药用植物, 单子叶, 茎直立, 叶线状披针形, 互生, 无叶柄(图1)。2013年经鳞茎播种后5叶及以上单株开始开花, 花单生于茎顶, 偶见顶部抽生2朵花, 花朵大, 初现花蕾绿色, 成熟花暗紫色, 花柄较长, 花弯曲下垂。花两性, 同被花, 花被未分化为萼片和花瓣(sepals and petals), 花被6, 2轮, 表面有黄褐色小方格花斑, 先端不卷曲; 花钟形, 花被片长2.5~3.2 cm, 内3片宽约10 mm, 外3片宽约6 mm; 单体雄蕊(stamen) 6枚, 2轮, 5枚雄蕊围绕雌蕊(pistil), 1枚雄蕊距离较远, 雄蕊由花药和花丝组成, 成熟花雄蕊长约为花被片的一半, 花丝黄绿色, 花药较长, 药隔明显; 合生雌蕊1枚, 柱头3裂, 裂片短, 毛状, 湿型, 表面有分泌物(exudate), 长0.5~1.2 mm, 子房上位, 花下位, 子房圆柱形, 紫色和黄绿色纵向相间。绿色花蕾中柱头高出雄蕊, 裂片外露, 当花被转为紫色开放后, 雄蕊花丝伸长, 将花药顶出, 使柱头低于花药, 包在5枚雄蕊中间, 既方便自交, 又保护柱头(图1)。田间观察到访花昆虫有黄蜂、蜜蜂、蝇类、蜘蛛等, 但数量极少。
 | 图1 漳县石川乡栽培暗紫贝母开花株形态及花被特征 A:驯化地生境Domesticated field surrounding; B:现蕾单株 Budding plant; C:开花前花器形态Floral organ shape before opening; D:柱头长出雄蕊 Longer stigma than the stamen; E:开花单株 Blooming plant; F:花被Perianth; G:开花Opening flower; H:柱头短于雄蕊Shorter stigma than the stamen.Fig.1 The flower shape and perianth characteristic for the F. unibracteata plants cultivated in Shichuan town of Zhang County |
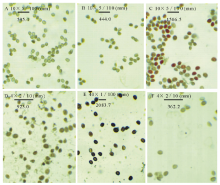
以花被开放当天为标准(0 d), 经对不同发育阶段的花朵花粉活力和柱头可受性测定显示(图2~4和表2), 暗紫贝母花朵在发育的不同时期其花粉粒总数、有活力花粉粒数均具有极显著差异(P< 0.01)。视野内有活力花粉粒数随着花朵的发育呈“ 正态分布” 趋势(图2)。当绿色花蕾长度达1.5 cm时, 即在花朵开放和花药开裂前2 d(-2 d)花粉已具有一定活力, 但活力很弱, 无色花粉量最大, 占总花粉量的83.7%(图2~3, 表2)。其柱头浸入联苯胺-过氧化氢液后有连续蓝色气泡产生, 显示柱头已具较强的可受性(图4和表2)。当花蕾长度增大到1.7 cm, 开花前1 d(-1 d), 弱活力花粉粒显著增多(P< 0.05), 花粉活力极显著增强(t=-3.660, P< 0.01), 无色花粉量极显著下降, 占总花粉量的47.1%(P< 0.01), 视野内仍未观察到强活力花粉粒, 但柱头周围蓝色气泡明显增多, 柱头可受性明显增强(图3和表3)。当花朵长度达2.5 cm, 花被变为暗紫色, 花被微开, 即为开花当天(0 d), 视野内花粉粒最多, 花粉活力最强, 平均花粉粒数达150粒以上, 有活力花粉粒在100粒以上, 均极显著高于其他发育阶段的水平(P< 0.001), 强活力花粉粒大量出现, 因花粉量增大, 导致无色花粉量有所增加, 但占总花粉量的比率下降为38.4%(图2~3), 此阶段柱头联苯胺-过氧化物酶活性也最强, 柱头可受性达到最高, 并维持至少3 d以上时间, 花粉活力最强持续时期与柱头可受性最强持续时期相一致, 但强活力花粉粒仅维持3 d时间(图3~4和表2)。开花后第1天(1 d)视野内花粉粒数量居第2位, 平均花粉粒数也在150粒以上。同时, 由于大量成熟花粉粒开始破裂, 内含物泄出, 导致统计的有活力花粉粒比率极显著下降(P< 0.01)。开花第2天(2 d)开始花粉粒数量极显著减少, 但强活力花粉粒比例仍维持在较高水平, 与前2 d差异不显著(P> 0.05)。开花后第3天(3 d)花粉量和活性均极显著下降(P< 0.01), 视野内强活力花粉粒消失, 仅观察到少量弱活性花粉粒, 并出现拖尾现象, 柱头反应气泡产生有所减少, 即柱头可受性呈减弱趋势(图3~4和表2), 至开花后第4天花瓣萎蔫闭合, 视野内花粉粒数量极少, 且全部失去活性, 柱头表现为较弱可受性, 局部已丧失可受性, 开花后第6天萎蔫花被开始脱落, 子房明显膨大。显然, 随着花朵发育进程的递进, 花粉量染色比率呈增大-减小趋势, 故无色花粉量随总花粉量和染色比率的变化而变化。
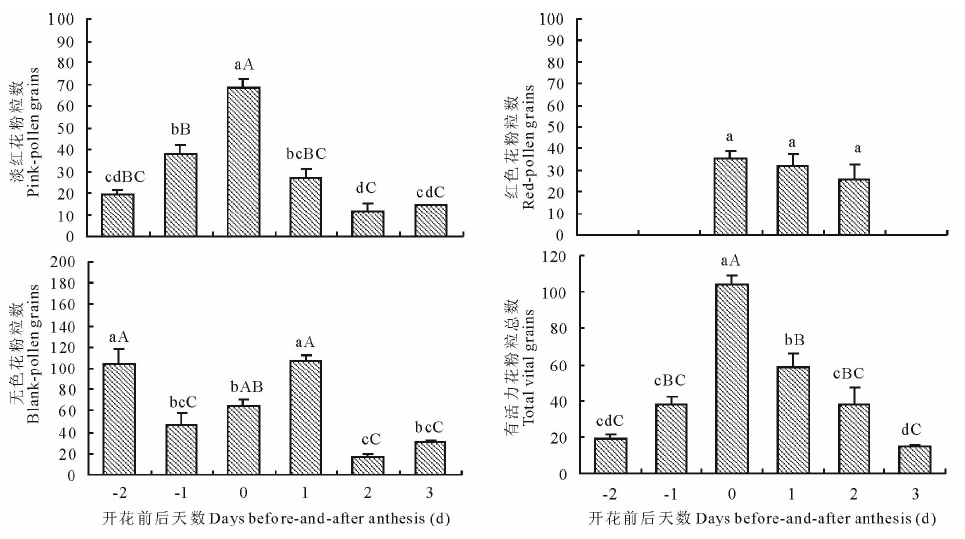 | 图2 漳县石川乡暗紫贝母开花过程中花粉粒数变化 图中数据为平均数± 标准误(n=15), 不同小写字母表示P< 0.05差异显著, 大写字母表示P< 0.01差异极显著。Fig.2 Changes of the pollination grains for F. unibracteata flowers during their development in Shichuan town of Zhang County The data in the Figure indicate means± SE(n=15). Different small letters mean significant difference at P< 0.05, capital letters mean great-significant difference at P< 0.01. |
 | 图3 漳县石川乡暗紫贝母花朵发育阶段花粉粒数量和活性变化动态 A:开花前2 d Two days before opening (-2 d); B:开花前1 d One day before opening (-1 d); C:开花当天 Opening day (0 d); D:开花后1 d One day after opening (1 d); E:开花后2 d Two days after opening (2 d); F:开花后3 d Three days after opening (3 d).下同The same below.4× , 10× , 40× :放大4倍、10倍和40倍Magnifies objects 4 times, 10 times and 40 times; 5/100 (mm), 5/100 (mm), 2/10 (mm): 比例尺及刻度单位Scale ruler and unit; 数字Numbers:清晰度 definition of objects.Fig.3 The numbers and viability of the pollination grains for F. unibracteata flowers from Shichuan town of Zhang County |
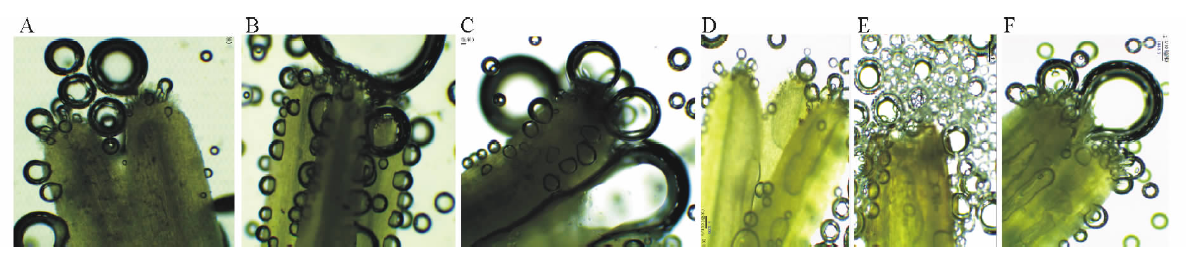 | 图4 漳县石川乡暗紫贝母花朵发育阶段柱头在联苯胺-过氧化氢液中反应变化动态(-2~3 d)Fig.4 Dynamic changes of the stigma receptivity reactive to the aniline-hydrogen peroxide for F. unibracteata flowers during development from Shichuan town of Zhang County |
| 表2 暗紫贝母花朵发育过程中花粉活力和柱头可受性的比较 Table 2 Comparison of pollen viability and stigma receptivity for F. unibracteata flowers during flowering process |
| 表3 暗紫贝母花朵发育阶段花冠形态变化 Table 3 Shape changes of the flower corolla during opening process for F. unibracteata |
经对绿色花蕾(-2 d)花药花粉粒和单花胚珠统计, 暗紫贝母成熟花蕾中每枚雄蕊含花粉粒约(1016± 340)粒, 单花含花粉(pollen)量约(6096± 2040)粒, 平均胚珠(ovule)数为160粒, P/O值为24.14~48.43, 平均37.5。按Cruden[9]的标准, P/O在31.9~396.0 范围, 繁育系统为兼性自交(facultative autogamy)。
经对20株暗紫贝母单花朵发育动态测定结果显示(表3), 暗紫贝母花朵开放过程中, 花冠显著伸长(F=4.706, P< 0.05), 极显著加宽(F=40.441, P< 0.01)。开花前2 d, 花冠直径已大于0.6 cm, 长度也在2.4 cm以上。从5月22日开花当天(0 d)至5月24日开花第3天(3 d), 花冠直径95%的置信区间为1.448~1.779 cm, 平均1.633 cm, 标准差0.455 cm, 变异系数27.84%。花冠长度95%的置信区间为2.657~2.860 cm, 平均2.758 cm, 标准差0.318 cm, 变异系数11.52%。开花后花冠加长但不显著(P> 0.05), 而花冠直径极显著加宽(P< 0.01), 显然暗紫贝母属于大花类型, 花朵营养生长与生殖生长并进。
依照Dafni[11]的评判标准(表1), 根据花朵直径对其杂交指数评判表明, 暗紫贝母成熟花朵的平均直径范围> 0.6 cm(表1), OCI记作3。暗紫贝母花药未开裂前柱头已具有较强的可受性, 花药开裂时间与柱头强可受性具有高度同步性(表2和图3~4), OCI记作0(表1); 在绿色花蕾中6枚雄蕊花药始终高于柱头, 当花被变紫并开放后, 即在开花当天(0 d), 雄蕊开始高出柱头, 6枚雄蕊花药与柱头间有空间分离(图1G~H), OCI记作1。综合上述结果, 暗紫贝母开放花朵OCI总和为4, 其繁育系统应属于部分自交亲和, 异交, 需要传粉者, 这与花粉量-胚珠比(P/O)的估算结果基本接近。
植物花的寿命随物种的不同而异, 产花粉能力低的花朵一般寿命较长, 柱头可受性可维持较长时间, 整个柱头活力期间, 花粉活性可被激活, 但单花间差异很大[17, 18]。胁迫生境下, 大量同时开花更易出现异交, 以改善后代的抗逆性[17]。圆叶风铃草(Campanula rotundifolia)高山居群长时间的柱头活力和高效率的传粉效果完全可以抵消由于活动昆虫少和频率低造成的不利影响[19]。青藏高原东部海拔影响暗紫贝母花大小和形态, 高海拔区花和花粉量均大, 但结种子数少[4]。本研究2013年在甘肃省漳县石川乡3100 m海拔范围内采用鳞茎栽培暗紫贝母, 2015年对开花植株观测发现, 鳞茎栽培3年的暗紫贝母种群开花期在5-6月, 单花寿命为6~8 d, 单花顶生弯曲, 偶见2花植株, 初现花蕾绿色, 后逐渐转为暗紫色, 花被2轮未分化, 离生雄蕊5枚紧围雌蕊, 1枚外围, 柱头3裂被毛, 有分泌物, 这种花部特征实现了其在高寒区极有限的适温范围内, 尽可能捕获光能, 短距离高效率为库容输送养料, 同时吸引昆虫, 并为其提供潜伏花内安全传粉的条件, 以弥补单株光合面积小, 传粉昆虫数量少的缺陷。暗紫贝母开花前后雌雄蕊空间错位, 开花当天花粉量最大, 有活力和强活力花粉粒率均最高。绿色花蕾柱头已具较强的可受性, 花药开裂时间与强柱头可受性同步, 其花粉活力的最高时期正是柱头可受的最佳时期, 但强活力花粉粒仅维持3 d时间, 而柱头可受性持续期在6 d以上, 后期由于花粉量急剧减少, 并有弱性花粉拖尾现象, 导致花粉活率高, 这与张晓军等[20]对平贝母的研究结果相似, 说明强活力花粉粒数指标更能反映花朵花粉活力状态。暗紫贝母花朵大小差异性大, 开花后花被加宽加长明显, 说明开花后单株有限的生长量集中在花部, 雄蕊高出柱头能把花药中的花粉聚拢在柱头上方, 雄蕊2轮着生还可增大访花昆虫花粉量携带部位面积, 提高高寒稀有传粉者的偶次访问中被同时带出花粉量大, 散布面广, 外力抖落花粉还易落于下方柱头可促进自花授粉, 花的弯曲下垂可保护花不被鸟食、阳光灼伤和风力损伤, 但却不利于花粉落于柱头实现自交。显然, 暗紫贝母的离生雌蕊与柱头空间错位可能与花弯曲下垂、花被不分离、柱头可受性持续期长、强活力花粉持续时间短联系在一起, 形成了适应自交、风媒和虫媒传粉异交的花部特征。
为了适应外界多面的环境, 植物形成了各种各样的繁育系统类型[8, 13]。有性繁殖是通过有性过程产生雌雄配子形成种子, 进而繁殖后代的一种繁殖方式, 有完整的个体发育周期[21]。自然界中, 很难见到绝对的异交或者绝对自交的植物类群, 大部分植物是二者兼有的混交模式[17]。自然授粉既包含自花授粉, 也包含异花授粉, 育种工作中一般根据异交率判断繁育类型[21]。张晓军等[20]研究表明, 平贝母(F. ussuriensis)繁育系统是以自花传粉为主, 虫媒异花传粉为辅的兼性自交。本研究结果显示, 川贝母基原植物暗紫贝母属于大花类型, 花朵直径为1.448~1.779 cm(> 0.6 cm), 花药开裂时间与柱头可受性具有同步性, 花药与柱头之间出现空间分离的现象, 其杂交指数(OCI)总和为4, 单花含花粉量约6000粒左右, 平均胚珠数为160, P/O值平均约37.5。综合评判, 暗紫贝母繁育系统应属于部分自交亲和, 异交, 需要传粉者。传粉媒介主要为风媒和虫媒, 田间观察到稀有访花昆虫有蜂类、蝇类、蜘蛛等, 这种异交和自交的交叉式繁殖方式传粉效率高。
致谢:漳县康荣中药材合作社康彦俊提供试验地和合作,中国科学院大学西北生态环境资源研究院博士生陈翔,兰州大学硕士生郭爱峰,甘肃农业大学本科生王军、李永明、杨锦强、何炳江、宋重昶参与田间播种。甘肃农业大学硕士生周传猛、杨慧珍、王小琴、梁伟、何媛丽、袁洪超、金彦博、郭一青,本科生倪吉星、年想红、李鹏、汤安参与田间试验,在此一并致谢。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|