


{kind=link}
{kind=link}
{kind=link}
套作对大豆苗期碳氮物质代谢的影响及其与抗倒伏性的关系
[任胜茂2, **  , 邓榆川
, 邓榆川2, ** , 文凤君2 , 刘明洁3 , 袁小琴4 , SajadHussain1 , 蒲全明2 , 刘卫国1, * , 杨文钰1, * ]
, 邓榆川, 文凤君, 杨文钰]
|
|


作者简介:任胜茂(1963-),男,四川南充人,高级农艺师。E-mail: 1802475325@qq.com。邓榆川(1989-),男,四川资阳人,研究实习员。E-mail: 515271439@qq.com。**共同第一作者These authors contributed equally to this work.
为从植株光形态建成的角度,阐明玉米-大豆套作模式下,大豆苗期倒伏发生的原因。在大豆单作和玉米-大豆套作两种种植模式下,以强耐阴大豆南豆12和弱耐阴大豆南032-4为试验材料,对大豆倒伏率、茎秆形态、叶片光合特性,茎秆和叶片碳氮代谢物质含量等进行调查、测定和分析。结果表明,由于套作受共生期内玉米的遮阴,大豆处于弱光环境,导致植株形态发生改变,光合速率降低,并发生倒伏,但两种大豆品种受荫蔽影响的程度不同。南豆12受荫蔽的影响较小,倒伏率显著低于南032-4,茎秆长粗比增加幅度和光合速率降低幅度也显著低于南032-4,相关分析表明,倒伏率与茎秆长粗比呈极显著正相关( r=0.946; P<0.01);与叶片光合速率呈显著负相关( r=-0.886; P<0.05);叶片光合速率与光合有效辐射呈极显著正相关( r=0.900; P<0.01),说明套作荫蔽降低了大豆冠层的光合有效辐射而导致大豆叶片光合速率降低是引起套作大豆形态改变,发生倒伏的重要原因;分析大豆叶片光合速率、茎秆碳氮比、叶片碳氮比和倒伏率发现,套作显著降低了茎秆和叶片碳氮比,降低幅度表现为南032-4显著高于南豆12,相关分析表明,叶片光合速率与茎秆和叶片碳氮比呈显著或极显著负相关( r=-0.871,-0.930; P<0.05),茎秆和叶片碳氮比与倒伏率呈极显著正相关( r=0.985,0.968; P<0.01),说明较高的碳氮比是南豆12具有较强抗倒伏能力的生理基础,使其能够在套作环境下维持较优的光形态特性,更适合于套作种植;分析大豆碳氮物质代谢可知,套作显著降低了大豆茎秆和叶片的碳氮代谢物质含量,不同品种间表现为南豆12显著高于南032-4。说明较高的碳氮代谢活性和光合产物运输能力是增强套作大豆抗倒伏能力的物质基础和代谢基础。以玉米-大豆带状套作种植为对象,探明了套作大豆植株倒伏与茎叶碳氮代谢物质转运的关系,为培育套作专用的耐阴抗倒伏大豆品种提供理论支持。
, DENG Yu-chuan, WEN Feng-jun, YANG Wen-yu
The purpose of this study was to clarify the role of photosynthetic capacity of soybean at the seedling stage in maize soybean intercropping. Our research studied the strongly shade tolerant soybean variety (Nandou 12) and weakly shade tolerant variety (Nan032-4), under soybean monoculture and maize soybean intercropping systems. The percentage of lodging,stem morphology, photosynthetic rate, carbon content and nitrogen metabolic activity of stems and leaves were measured. Compared with monoculture soybean, the stem length:diameter ratio was significantly increased by maize shading, but leaf photosynthetic rate and the lodging percentage were decreased. However, the degree of response differed between varieties. In the intercropping system, the stem morphology changes and increase in lodging were relatively small in the shade tolerant variety Nandou 12, and greater in the intolerant variety, Nan032-4. Lodging percentage was highly significantly correlated ( r=0.946; P<0.01) with the stem length:diameter ratio, and negatively correlated with the photosynthetic rate ( r=-0.886; P<0.05).Meanwhile, the photosynthetic rate was highly significantly positively correlated with the photosynthetically active radiation ( r=0.900; P<0.01). The results show that shading by maize reduced the photosynthetically active radiation and photosynthetic rate, which in turn triggered plant height increase and stem diameter decrease responses, leading to increased plant lodging. Compared with soybean monoculture, the carbon:nitrogen ratio of intercropped soybean was significantly decreased by maize shading, but again Nandou 12 was significantly less affected than Nan032-4. Leaf photosynthetic rate was highly significantly and negatively correlated with the carbon:nitrogen ratios of leaf and stem ( r=-0.871 and -0.930, respectively; P<0.05). The leaf and stem carbon:nitrogen ratios were highly significantly ( r=0.985 and 0.968, respectively; P<0.01) and positively correlated with lodging percentage. These findings show that a high carbon:nitrogen ratio in soybean stem and leaf tissues provides a physiological basis for maintenance of higher lodging resistance and the variety with this trait (Nandou 12) was more suitable for intercrop planting than Nan032-4. Compared with soybean grown in monoculture, the content of carbon and nitrogen and photosynthetic activity and substrate export of stem and leaf tissue was significantly decreased when shaded by maize in the intercrop system. Soybean monoculture and maize soybean intercropping systems were used as in this study to explore the relationship between plant lodging, and carbon and nitrogen metabolism to provide a theoretical basis for development of appropriate fertilization management and cultivation practices to promote lodging resistance and identify varieties to lodging in intercropping systems.
近年来, 随着我国耕地面积的不断减少, 土壤肥力的逐年降低, 农田生态环境的恶化加剧, 传统套作种植模式以其“ 保地增肥、增加产出” 的优势被人们重新重视, 以玉米(Zea mays)-大豆(Glycine max)带状套作模式最为突出, 被农业部连续多年认定为农业主推技术。该模式充分利用了豆科与禾本科套作体系高效的氮素利用、合理的空间搭配和更优的边际效益等特点, 在保证玉米不减产的前提下增加一季大豆的产量, 从而提高土地产出率, 具有省工、节本、高产高效和利于产业化等优点[1, 2]; 对改善农田生态环境, 改良农田土壤质量, 提高土地利用率, 解决我国人地资源矛盾等方面起到了重要作用。
作物的生长发育与气候环境密切相关[3, 4], 随着玉米-大豆带状套作种植模式下大豆局部小气候研究的不断深入, 发现光照是影响套作大豆生长发育的关键因子。研究表明, 套作大豆受其共生期内玉米的遮阴而处于弱光照强度和低红光/远红光环境, 导致大豆叶面积指数下降, 比叶重降低, 茎秆纤细, 节间过长, 在苗期极易发生倒伏和藤蔓化生长[5, 6, 7, 8]。而影响套作大豆倒伏的因子较多, 以品种为主, 气候环境和栽培措施等也会引发或加重倒伏[5, 6, 7, 8, 9]。研究表明, 植株光形态建成是影响大豆倒伏的最主要因素, 而植株对光合产物的高效积累与合理分配是优良形态形成的基础, 对提高大豆抗倒伏能力起着至关重要的作用[10]。碳氮代谢作为植物体内最为重要的两大代谢途径, 其代谢的强弱直接影响着光合产物的积累与分配[11], 而较高的碳氮代谢酶活性对植株的抗倒伏性具有明显的促进作用[12]。研究表明, 碳水化合物是作物生长发育过程中的重要组成部分, 其含量的变化能够体现植物体内碳水化合物的合成与运输情况[13]。同样, 氮素对光合产物的积累和分配也起着重要的调节作用[14]。碳氮比不仅能反映植株碳氮化合物代谢能力, 还能体现植物的氮素利用效率, 在作物整个生育期间较高的碳氮比能够促进植株生长发育, 优化植株形态[15]。
茎秆和叶片是套作大豆苗期“ 源-库-流” 的重要组成部分, 是大豆光形态建成的关键[16]。而前人对大豆植株倒伏性的研究大多是基于植株地上部、地下部或是针对某一器官的单一研究, 且集中于氮素调节和资源筛选方面[17]。并没有分析套作大豆苗期的弱光胁迫是如何影响光合产物的积累和茎叶碳氮代谢物质运输, 使得大豆植株的形态和倒伏性等方面产生了差异[18]。因此, 选取耐阴、抗倒伏性具有显著差异的两个大豆品种, 在单作和套作两种种植模式下, 分析比较茎秆形态、叶片光合、碳氮物质代谢等性状指标之间的差异, 研究套作荫蔽对大豆光合产物积累的影响、茎叶碳氮代谢物质的转移运输规律及其与植株倒伏性的关系, 以明确套作大豆耐阴抗倒伏的碳氮物质代谢基础。
试验于2015年在四川农业大学雅安校区教学农场进行(29° 59' N, 103° 00' E), 年积温5071.5 ℃, 年降水1732 mm, 年平均气温16.2 ℃, 年无霜期304 d, 年平均日照时数1039 h。试验地为重壤地, 有机质29.86 g· kg-1、全氮716.43 mg· kg-1、速效氮114.88 mg· kg-1、碱解氮62.29 mg· kg-1、速效磷36.85 mg· kg-1、速效钾136.61 mg· kg-1, pH 7.4。供试大豆材料为强耐阴的南豆12和弱耐阴的南032-4[5]。玉米品种为正红505。
试验采用裂区设计, 主区为玉米-大豆带状套作和大豆单作2种种植模式, 副区为2种大豆品种, 试验重复3次。套作模式中, 按照文献[18]采用宽窄行种植, 带宽2 m, 宽行1.6 m, 窄行0.4 m, 玉米种植于窄行, 大豆种植于宽行, 行距均为0.4 m, 每带种2行玉米和2行大豆, 每个小区种6带, 带长12 m, 小区面积为72 m2; 单作模式中, 大豆采用等行距种植, 行距为0.5 m, 长6 m, 每小区种6行, 小区面积为18 m2。
玉米于2015年4月2日育苗, 4月9日移栽, 穴距0.2 m, 每穴1株; 大豆于2015年6月18日播种于玉米宽行内, 穴距0.1 m, 与玉米行相隔0.6 m, 6月26日间苗, 每穴定苗1株。施肥和其他管理措施同大田生产。
1.3.1 倒伏调查 实际倒伏率 (actual lodging percentage, ALP):套作大豆出苗后35 d小区内实际倒伏植株占该小区全部植株的比率, 以主茎与地面的夹角≤ 30° 认定其为倒伏。植株倒伏程度采用周蓉等[19]的评价方法。
1.3.2 光合特性 同一时期按小区选取长势一致的植株5株, 用于测定植株叶片光合特征参数。
光合有效辐射的测定方法:采用光量子传感器Li-191SA(LI-COR, USA)在晴朗无风的上午11:00-12:00测定玉米冠层上方入射光合有效辐射和套作大豆冠层光合有效辐射, 重复测定5次, 透光率的计算方法:透射光与入射光的比值。
光合特征参数的测定方法:利用光合测定仪LI-6000(LI-COR, USA)测定由上至下第3片复叶的光合速率(photosynthetic rate, Pn)、气孔导度(stomatal conductance, Gs)、蒸腾速率(transpiration rate, Tr)、胞间二氧化碳浓度(intercellular CO2 concentration, Ci)、光合有效辐射(photosynthetically active radiation, PAR)等光合特性参数。
叶绿素的测定方法:迅速截取第3片复叶经液氮处理后保存于冰盒中用于叶绿素含量的测定, 其测定参照Sims等[20]的方法。
叶面积的测定方法:采用打孔法[10]测定。
1.3.3 茎秆形态 同一时期按小区选取长势一致的植株10株测定茎秆形态。
测定方法:用刻度尺测量[10]从地上部分开始至主茎顶端生长点的实际长度为株高; 用游标卡尺测量[10]植株中部节间直径取平均值为茎粗。
1.3.4 碳氮代谢物质 同一时期按小区选取长势一致的植株10株迅速截取植株茎秆和叶片分别装入纸袋, 在105 ℃下杀青0.5 h并于75 ℃下烘干至恒重后, 粉碎过0.3 mm (60目)筛, 测定碳氮代谢物质含量。
测定方法:淀粉含量的测定参考张志良等[21]的方法; 用考马斯亮蓝法[21]测定可溶性蛋白含量; 用FOSS-SOX 500型脂肪测定仪测定粗脂肪含量; 用CE-440型元素分析仪测定总碳; 总氮测定参考王海艳等[22]的方法, 均重复测定3次。
碳氮比的计算方法:总碳与总氮的比值。
用Microsoft Excel 2007软件整理数据, 用Excel绘图, 用SPSS 17.0软件分析数据, 采用Duncan氏新复极差(SSR)法进行显著性检验。
2.1.1 倒伏率 套作大豆苗期倒伏严重(图1a), 不同品种大豆倒伏发生有显著差异, 表现为南032-4> 南豆12; 根据周蓉等[19]的评价方法, 南032-4的倒伏率达80.37%, 属于严重倒伏, 南豆12为47.04%, 属中度倒伏。说明在套作模式下南豆12较南032-4具有较强的抗倒伏能力。
2.1.2 茎秆长粗比 由图1b可知, 套作显著增加了大豆茎秆的长粗比, 但不同品种大豆受套作荫蔽影响的程度不同。与单作相比, 套作模式下南032-4长粗比增加了1.41倍, 而南豆12增加了1.26倍, 差异极显著(P< 0.05)。说明强耐阴大豆南豆12受套作荫蔽的影响程度相对较小, 在套作下表现出较优的茎秆特性。
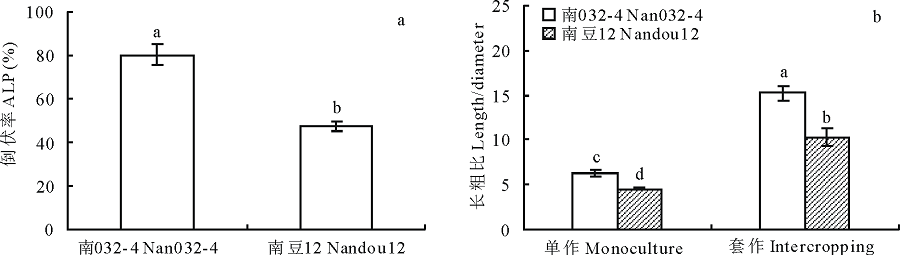 | 图1 套作对大豆倒伏率和茎秆长粗比的影响不同字母表示差异达0.05显著水平。下同。Fig.1 Effects of different planting patterns on actual lodging percentage and length/diameter of soybeanThe different letters mean significant difference at 0.05 level. The same below. |
2.2.1 叶面积 由表1可知, 套作显著降低了大豆的叶面积, 但不同品种大豆受套作荫蔽影响的程度不同。与单作相比, 套作模式下南032-4降低了27.41%, 而南豆12仅降低20.00%, 差异极显著(P< 0.01)。方差分析表明, 大豆品种与种植模式间互作对叶面积的影响极显著(P< 0.01)。相关性分析表明, 大豆叶面积与光合速率呈极显著正相关(r=0.784, P< 0.01)(表2)。
| 表1 不同种植模式对大豆叶片光合特性的影响 Table 1 Effects of different planting patterns on photosynthetic characteristics of soybean leaves |
2.2.2 叶绿素含量 由表1可知, 套作显著降低了大豆叶片的叶绿素含量, 但不同品种大豆受套作荫蔽影响的程度不同。与单作相比, 套作模式下南032-4叶绿素含量降低了34.38%, 而南豆12仅降低11.23%, 差异极显著(P< 0.01)。方差分析表明大豆品种与种植模式间互作对叶绿素含量的影响极显著(P< 0.01)。相关性分析表明, 大豆叶绿素含量与光合速率呈极显著正相关(r=0.878, P< 0.01)(表2)。
2.2.3 光合特性 由表1可知, 套作荫蔽降低了大豆冠层光合有效辐射, 导致大豆叶片光合速率、气孔导度和蒸腾速率显著降低, 胞间二氧化碳浓度显著提高(P< 0.01), 而品种间差异极显著(P< 0.01), 大豆品种和种植模式间互作对叶片光合特性的影响极显著(P< 0.01)。与单作相比, 套作模式下的透光率为53.88%, 南032-4和南豆12的光合速率分别降低了37.29%和36.18%, 说明套作模式下强耐阴品种南豆12能保持较高水平的光合速率, 具有较强的荫蔽适应性。胞间二氧化碳浓度与光合速率的变化趋势相反, 而气孔导度和蒸腾速率与光合速率的变化趋势相同, 相关分析表明, 气孔导度、蒸腾速率、光合有效辐射与光合速率存在极显著的正相关关系, 相关系数分别为0.916、0.968、0.900(P< 0.01, 表2), 与胞间二氧化碳呈极显著负相关, 相关系数为-0.934(P< 0.01)。说明套作模式下, 大豆叶片蒸腾速率降低, 气孔导度变小限制了二氧化碳的扩散, 从而导致光合速率降低。
2.3.1 淀粉 大豆叶片同化的光合产物主要以积累淀粉为主, 其含量变化是反映植株光合能力强弱和体内碳积累状况的重要指标[23]。由图2可知, 套作显著降低大豆叶片的淀粉含量, 但受影响的程度因大豆品种而异, 而不同种植模式下均表现为南豆12显著高于南032-4。与单作相比, 套作模式下南032-4叶片淀粉含量降低了21.15%, 而南豆12仅降低18.07%, 两品种间的差异显著(P< 0.05)。方差分析表明大豆品种与种植模式间互作对大豆叶片淀粉含量的影响显著(P< 0.05)。说明大豆叶片淀粉的积累是由种植模式和品种共同调控作用的, 而强耐阴的南豆12表现出更高的淀粉积累能力, 为茎秆抗倒伏形态建成提供了充分的物质保障。
| 表2 大豆叶片光合指标的相关性分析 Table 2 Correlation coefficients among photosynthesis indexes of soybean leaves |
2.3.2 粗脂肪 脂肪不仅在维持植物生长、发育过程中起着重要作用, 而且还在适应外界环境变化中扮演着重要的角色[24]。由图2可知, 套作显著降低大豆叶片的粗脂肪含量, 但受影响的程度因大豆品种而不同, 在不同种植模式下均表现为南豆12显著高于南032-4。与单作相比, 套作模式下南032-4叶片粗脂肪含量降低了18.71%, 而南豆12仅降低16.67%, 两品种间的差异显著(P< 0.05)。方差分析表明大豆品种与种植模式互作对大豆叶片粗脂肪含量的影响显著(P< 0.01)。说明套作模式下, 强耐阴的南豆12具有更强的荫蔽适应能力, 更适合于套作种植。
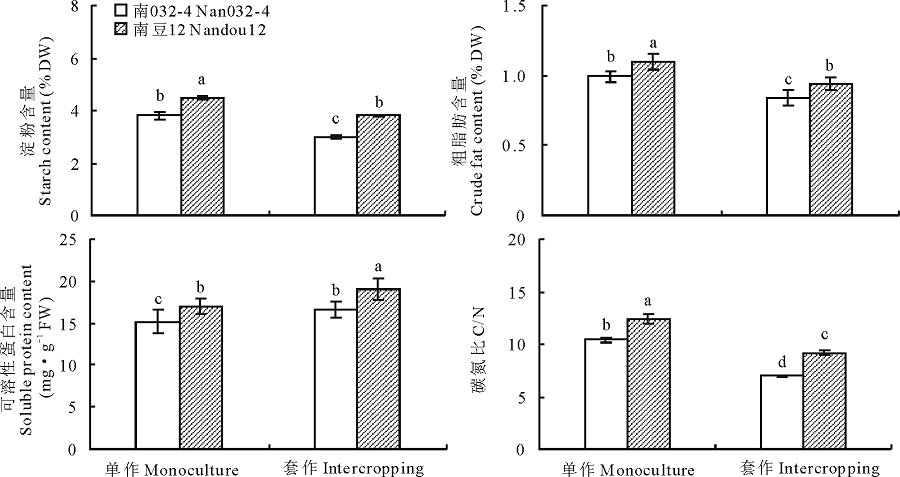 | 图2 不同种植模式对大豆叶片中碳氮代谢的影响Fig.2 Effects of different planting patterns on carbon and nitrogen metabolism of soybean leaves |
2.3.3 可溶性蛋白 可溶性蛋白是植物所有蛋白质组分中最活跃的一部分, 包括各种酶原、酶分子和代谢调节物, 其含量是反映作物的氮素营养、氮素同化作用与再利用状况、叶片功能及抗性等的重要指标[25]。由图2可知, 套作显著提高了大豆叶片的可溶性蛋白的含量, 但受影响的程度因大豆品种而不同, 在不同种植模式下均表现为南豆12显著高于南032-4。与单作相比, 套作模式下南032-4叶片可溶性蛋白含量提高了9.15%, 而南豆12提高12.75%, 两品种间的差异显著(P< 0.05)。方差分析表明大豆品种与种植模式互作对大豆叶片可溶性蛋白含量的影响显著(P< 0.01)。
2.3.4 碳氮比 碳氮比不仅反映植株碳氮代谢的强弱, 对调节植株生长也起着重要作用[25]。由图2可知, 套作显著降低大豆叶片的碳氮比, 但受影响的程度因大豆品种而不同, 在不同种植模式下均表现为南豆12显著高于南032-4。与单作相比, 套作模式下南032-4叶片碳氮比降低了33.21%, 而南豆12仅降低26.16%, 两品种间的差异显著(P< 0.05)。方差分析表明大豆品种与种植模式互作对大豆叶片碳氮比的影响极显著(P< 0.01)。
2.4.1 淀粉 由图3可知, 套作显著降低大豆茎秆的淀粉含量, 但受影响的程度因大豆品种而异, 在不同种植模式下均表现为南豆12显著高于南032-4。与单作相比, 套作模式下南032-4茎秆淀粉含量降低21.60%, 南豆12降低19.11%, 差异显著。方差分析表明大豆品种与种植模式互作对大豆茎秆淀粉含量的影响极显著(P< 0.01)。
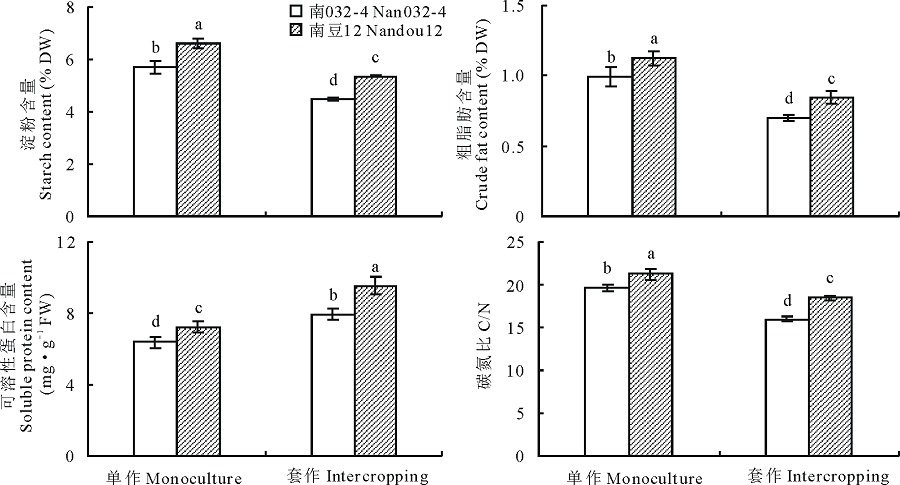 | 图3 套作对大豆茎秆中碳氮代谢的影响Fig.3 Effects of different planting patterns on carbon and nitrogen metabolism of soybean stem |
2.4.2 粗脂肪 由图3可知, 套作显著降低大豆茎秆的粗脂肪含量, 但受影响的程度因大豆品种而不同, 在不同种植模式下均表现为南豆12显著高于南032-4。与单作相比, 套作模式下南032-4茎秆粗脂肪含量降低了29.19%, 而南豆12仅降低24.74%, 两品种间的差异显著(P< 0.05)。方差分析表明大豆品种与种植模式互作对大豆茎秆粗脂肪含量的影响极显著(P< 0.01)。
2.4.3 可溶性蛋白 由图3可知, 套作显著提高了大豆茎秆的可溶性蛋白的含量, 但受影响的程度因大豆品种而不同, 在不同种植模式下均表现为南豆12显著高于南032-4。与单作相比, 套作模式下南032-4茎秆可溶性蛋白含量提高了24.62%, 而南豆12提高31.68%, 两品种间的差异显著(P< 0.05)。方差分析表明, 大豆品种与种植模式互作对大豆茎秆可溶性蛋白含量的影响极显著(P< 0.01)。
2.4.4 碳氮比 由图3可知, 套作显著降低大豆茎秆的碳氮比, 但受影响的程度因大豆品种而异, 在不同种植模式下均表现为南豆12显著高于南032-4。与单作相比, 套作模式下南032-4茎秆碳氮比降低了18.48%, 而南豆12仅降低12.90%, 两品种间的差异显著(P< 0.05)。方差分析表明, 大豆品种与种植模式互作对大豆茎秆碳氮比的影响极显著(P< 0.01)。
对套作大豆茎秆形态、光合特性、田间倒伏率、碳氮代谢物质含量进行相关性分析, 结果表明(表3), 大豆田间倒伏率与茎秆长粗比呈极显著正相关(r=0.946, P< 0.01), 与叶面积呈极显著负相关(r=-0.874, P< 0.01), 说明较优的茎秆形态能够提高作物的抗倒伏能力; 大豆叶片光合速率与田间实际倒伏率呈显著负相关(r=-0.886, P< 0.05), 与大豆冠层光合有效辐射呈极显著正相关(r=0.900, P< 0.01), 说明套作遮阴降低了大豆冠层的光合有效辐射而导致大豆叶片光合速率的降低是影响套作大豆苗期倒伏的关键因素; 叶片和茎秆碳氮比与倒伏率、叶片光合速率、茎秆长粗比均呈显著或极显著相关性。说明植株碳氮代谢是光合产物的积累和运输的关键, 而在套作下, 植株正常的碳氮代谢过程受阻, 叶片光合产物的积累量降低, 代谢物质含量也降低; 茎秆节间氮含量增加, 厚壁细胞的物质充实性降低, 从而改变植株的光合形态建成, 影响植株的倒伏。
| 表3 大豆碳氮代谢与倒伏率、光合速率相关性分析 Table 3 Correlation analysis of carbon and nitrogen metabolism, actual lodging percentage and photosynthetic rate in different planting patterns |
光照主要以环境信号的形式调节植物生长、发育和形态建成[26]。随着玉米-大豆带状套作模式下农田小气候研究的不断深入, 发现在玉米-大豆共生期内, 高位作物玉米的遮阴, 不仅降低了大豆冠层光合有效辐射强度, 还由于玉米对自然光的选择性吸收降低了红光与远红光的比例[5, 6, 7], 使大豆发生避阴性反应, 茎秆变细, 节间伸长, 容易发生倒伏。本试验进一步表明, 套作模式下植株茎秆变细, 株高增加, 叶面积降低, 叶绿素含量降低, 田间倒伏严重。刘婷等[10]在研究遮阴对大豆光合特性和倒伏性的影响中表明, 遮阴降低了大豆叶片的光合速率(Pn), 增加了倒伏的发生, 而导致大豆叶片光合速率降低的主要原因是胞间二氧化碳(Ci)浓度过高超过了叶片正常光合作用的需求量, 气孔导度(Gs)的降低又极大地限制了二氧化碳的扩散, 从而导致光合速率降低。研究还表明, 大豆的抗倒性不仅与基因型密切相关, 还与植株叶片光合特性、光合产物积累分配代谢等有关[10, 16, 27]。基于前人的研究结论, 对不同种植模式下叶片光合参数进行测定和田间倒伏调查。进一步研究发现, 大豆苗期叶片光合速率与倒伏率呈显著负相关(r=-0.886, P< 0.01)。荫蔽下倒伏发生严重可能是大豆受弱光胁迫, 光合产物的积累分配改变, 正常的植株生理代谢受阻, 导致植株形态建成改变[10]。而2种大豆的光合速率均降低, 但强耐阴性品种南豆12降低程度较不耐阴性品种南023-4低, 表现出较优的抗倒性和较强的光合特性, 适合套作种植。
碳氮代谢是作物生长发育过程中最基本的代谢途径, 两大代谢相互协调共同调节作物的生长发育进程, 影响作物的产量形成[28]。而影响作物碳氮代谢的因素很多, 主要有植株基因型、生长发育阶段、器官以及生长环境条件等[15], 其中光照对植物碳氮代谢起着至关重要的作用。适宜的光照能保证光合作用的正常进行, 合成植物生长所必需的碳源, 为氮代谢提供能量[16], 而在荫蔽胁迫下, 作物的碳氮代谢活性将会受到抑制[29, 30]。关义新等[31]的研究表明, 较强的光照条件下玉米植株具有较高的碳氮同化能力和代谢物含量。王竹等[6]的研究表明, 当套作大豆荫蔽程度得到缓解时, 植株的碳氮含量和相关酶的活性均显著增加。本试验中, 套作荫蔽显著降低了大豆植株的碳氮代谢活性, 表现为碳氮代谢物质含量显著低于单作。与南032-4相比, 强耐阴性大豆南豆12受套作遮阴影响较小, 其叶片光合效率, 淀粉含量、可溶性蛋白均较高, 表现出较强的光合性能; 其茎秆中淀粉含量高, 碳氮比值高, 促进厚壁细胞的物质充实, 增加纤维素和木质素含量, 从而促进茎秆的形态建成, 提高茎秆的机械强度, 增加大豆的抗倒伏能力[17, 31]。说明较高的碳氮含量是大豆具有较强抗倒伏能力的物质基础, 使其能够在套作环境下维持较优的形态综合性状, 更适合于套作种植。
对于苗期大豆而言, 库主要是茎秆, 不仅是营养的贮存中心, 还是叶片光合产物的运输通道[16]。淀粉在植物体内与碳水化合物互相转化, 是植物碳代谢的主要产物之一, 其含量的变化与碳水化合物形成、转运和利用情况有关, 反映了同化物从源端叶片到库端茎秆的供应情况和运输、利用率, 其含量的高低直接与光合作用相关[16, 32]。可溶性蛋白质是植物体内的主要生理代谢物质, 在植株氮素代谢中起着代谢库的作用[33]。本研究中套作荫蔽显著降低了大豆的碳氮代谢活性和物质转移能力, 这与刘沁林等[16]的研究相一致。而强耐阴性的南豆12表现出较强的碳氮代谢能力, 可能是由于茎秆具有较强的可塑性, 对光合产物的分配起到了一定的调节作用。
在玉米-大豆带状套作种植下, 套作大豆苗期茎秆长粗比、植株倒伏率、叶片光合特性、茎叶碳氮代谢物含量及碳氮比与单作大豆相比差异显著, 除可溶性蛋白含量和茎秆长粗比高于单作, 其余指标均低于单作。通过相关性分析, 套作大豆倒伏率与茎叶碳氮代谢物质含量相关性均达到极显著水平(P< 0.01), 其中, 与叶面积、淀粉含量、粗脂肪含量、可溶性蛋白呈极显著负相关, 与碳氮比呈极显著正相关。特别是叶片碳氮比, 相关系数达到0.985。因此, 套作大豆苗期茎叶碳氮代谢与抗倒伏性密切相关。本研究为净套作大豆合理施肥管理、研究光环境对大豆光合形态建成和培育套作专用的耐阴抗倒大豆提供理论支持。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|