


{kind=link}
{kind=link}
{kind=link}
{kind=link}
二倍体菘蓝开花习性及传粉特性研究
[张金霞1  , 陈垣
, 陈垣1, 2, * , 郭凤霞1, * , 王引权3 , 周盛茂4 , 肖生伟5 ]
, 陈垣, 郭凤霞, 王引权|
|


作者简介:张金霞(1992-), 女,甘肃陇西人,在读硕士。E-mail: 1793545841@qq.com
菘蓝是药材板蓝根和大青叶的基原植物,依赖种子繁殖。通过田间观测和人工授粉实验,对民乐县栽培菘蓝开花时序和传粉生物学系统进行研究,旨在为其种子繁育和杂种优势利用提供科学和技术依据。研究表明,菘蓝为无限花序,单株表现异向开花习性,花序从植株顶部向下,单花从花序基部向上依次开放。花两性,雄蕊6,4枚长,2枚短,花萼和花瓣均为4,花萼绿色,花瓣黄色,子房上位,1室。单花寿命6~7 d,异常气候可缩短至3~4 d,群体开花期30 d左右。即将开放的花蕾柱头已成熟,花瓣刚露出花萼后开花当天,花粉鲜黄而散,末花期子房膨大,花粉干枯。短角果结种子0~2粒。自然结籽率95.0%,不去雄和去雄套纸袋均不结实,同株异花去雄授粉结籽率仅4.0%,去雄套纸袋异株异花授粉结籽率72.0%,去雄不套袋结籽率53.3%,自交不亲和指数为0.96~1.00。不去雄套网袋结籽率5.0%,去雄后提高至13.7%,访花者主要有食蚜蝇、黑蜂等。说明菘蓝具有较强的自交不亲和性,不存在无融合生殖现象,属典型的异花授粉类型,需传粉者,传粉媒介以虫媒为主,风媒为辅。
Isatis indigotica is the primary ingredient of traditional Chinese drugs ‘Banlangen’ and ‘Daqingye’, and its cultivation relies on seed propagation. This research examined the flowering habit and pollination biology of I. indigotica cultivated in Minle county. The work included field observation and an artificial pollination experiment, in order to provide scientific and technical information to support seed production and heterosis utilization in this crop. It was found that the plant had an infinite inflorescence consisting of an upright main stem bearing raceme-like branches with new branches above as older matured branches below and 10-20 individual flowers per branch opening in an acropetal sequence over a flowering period of some 30 days. Flowers are bisexual with 6 stamens, four of which are long and two of which are short. There are 4 green calyces, 4 yellow petals with ovary superior, and 1 locule. The flower lifespan was 6-7 days but was shortened to 3-4 days under adverse climate conditions. The stigma matures just prior to the flowers opening. When the petals are just emerging from the calyx tube, bright fresh yellow pollen grains become visible and are dispersed by the flowering day. Anthers then begin to wither, and the ovary expands at the end of the flowering stage, growing a short pod with 0-2 seeds. Seed set was 95.0% for plants under natural conditions. Under covered conditions with paper bags, seed set was very low (approximately 4%) both with and without castration, but was 72.0% when plants were cross-pollinated after castration. When plants were cross-pollinated after castration, but the inflorescence was unbagged, seed set was 53.3%. The self-incompatibility index was assessed as 0.96-1.00. For bagged racemes, seed set was 5.0% without castration and increased to 13.7% after castration. The most common flower pollinator visitors were Episyrphus balteatus and Apis mellifer but various other insect species also visited. These results indicate that I. indigotica has strong self-incompatibility without apomixis, and can be considered a typical cross-pollinating plants needing pollinators, pollinated mainly by insects but supplemented by wind.
菘蓝(Isatis indigotica)为十字花科(Cruciferae), 菘蓝属(Isatis)二年生草本药用植物[1]。药材板蓝根(Radix isatidis)为菘蓝的干燥根, 秋季采挖, 除去泥沙, 晒干, 具有清热解毒、凉血利咽等功效, 用于瘟疫时毒, 发热咽痛, 温毒发斑, 痄腮, 烂喉丹痧, 大头瘟疫, 丹毒, 痈肿等症。大青叶(Folium isatidis)为菘蓝的干燥叶, 夏秋二季分2~3次采收, 除去杂质, 晒干, 具有清热解毒, 凉血消斑功效, 用于温度高热、神昏、发斑发疹、痄腮、喉痹、丹毒、痈肿等症[2]。板蓝根还具有抗炎、解热、提高免疫力和防治严重急性呼吸综合征(severe acute respiratory syndrome, SARS)[3]等流感病毒作用, 临床上被用于预防和治疗咽炎、扁桃体炎、肝炎、角膜炎等感染性疾病[4]。随着动物人畜共患病的传播流行, 板蓝根和大青叶需求量随之增加。然而, 菘蓝野生资源稀少, 板蓝根和大青叶药源主要依赖栽培品。甘肃省河西地区民乐县一带是我国板蓝根和大青叶的主要道地产区, 2012年被农业部和中国特产之乡推荐暨宣传活动组委会授予“ 中国板蓝根之乡” 。
种子播种栽培是菘蓝生产的主要方式[5]。作为板蓝根和大青叶的基原植物二倍体菘蓝, 在我国有着广泛的栽培[6], 但由于长期以来采用种子繁殖而不加以选择, 加上种子商品流通, 导致各地种植的品种混杂退化, 造成药材产量和品质参差不齐, 以此原料炮制的大青叶和板蓝根饮片及复方中成药品的疗效稳定性减弱。虽然部分产地已制定并实行菘蓝规范化种植和加工管理, 但这不能从根本上解决种源“ 质” 的问题。中药的品质是由遗传特性及生长环境决定, 其中遗传因素是决定中药材品质的内因, 环境是外因, 其影响程度与药材种类有关[7]。植物的繁育是植物内在遗传特性和外界环境相互作用的结果, 是种群有性生殖的纽带, 对植物的遗传变异和进化过程均有着重要的意义[8]。菘蓝品种随着种植年限的延长, 混杂退化问题更为突出, 严重影响药材板蓝根和大青叶的产量和内在品质[5]。王恩军等[9]对民乐县栽培菘蓝研究发现, 菘蓝种质资源具有丰富的遗传多样性, 主要农艺性状变异丰富, 变异系数(coefficient of variation, CV)为 9.83%~69.82%, 大青叶和板蓝根产量的CV均大于 50%。植物的花被特征和传粉生物学是反映植物有性生殖能力的重要依据[10, 11, 12, 13]。从植物开花习性入手, 详细研究其花部结构特征, 传粉媒介和传粉者在花间的运动样式、自交亲和程度以及交配系统才能从根本上了解其繁殖特性[14]。有关药用植物传粉特性的研究广泛而深入, 陈垣等[15]研究探明了暗紫贝母(Fritillaria unibracteata)的花器官特征, 郭凤霞等[16]进一步研究探明了其传粉生物学, 这些研究奠定了川贝母基原植物新品种选育的理论基础。尽管对菘蓝有性繁殖特性的研究也有少量报道, 叶青[5]研究了菘蓝生物学特性, 随后高延辉等[6, 17, 18]研究发现菘蓝花期自交不亲和性较强, 而蕾期自交亲和性较高, 并筛选了克服其自交不亲和性的化学试剂。这些研究对了解菘蓝生物学特性、自交不亲和性及其利用均具有重要参考价值, 但研究区与我国板蓝根主要道地产区民乐县地理及生态环境差异很大, 自交不亲和性的研究也仅是对吉林长春市生长2年的菘蓝从人工自花授粉、天然套袋自交和人工去雄异交处理后, 实验室花粉培养萌发及柱头显微镜检判断的结论, 由于植物的传粉特性不仅涉及自交不亲和性, 还包括传粉媒介和结籽率等, 并受基因型和外界环境因子的综合影响, 仅采用简单处理后在实验室培养条件下难以全面揭示其传粉特性, 去雄和不去雄花蕾在套网袋和纸袋条件下的结籽率和结实率均可反映其风媒和虫媒传粉的实证, 也可揭示有无孤雌生殖现象, 去雄后不套袋还可反映出自然异花授粉潜力, 但目前有关这些研究尚未见报道, 至今菘蓝种子繁殖操作规程也未有颁布, 生产上种子繁殖措施仍处于盲目阶段。因此, 在菘蓝道地产区民乐县自然环境条件下, 采用多种处理系统研究菘蓝开花习性和传粉特性具有重要意义, 可为菘蓝新品种选育和种子标准化生产提供科学和技术依据, 促进菘蓝种质资源的可持续化发展。
试验区位于甘肃省张掖市民乐县, 地处张掖市东南部, 祁连山北麓, 河西走廊中段, 地势南高北低, 地形分山地和倾斜高平原两大类, 属于温带大陆性荒漠草原气候, 海拔由北向南逐步从1589 m上升为5027 m, 年均降水量351 mm, 年均气温0~7.6 ℃, 无霜期140 d。民乐县地域辽阔, 气候冷凉, 土壤肥沃, 光照充足, 适宜菘蓝生长。当地生产的板蓝根色白、粉性足、根条粗壮, 总灰分含量低[19], 浸出物含量高, 品质优良, 被授予“ 中国板蓝根之乡” [19, 20, 21]。
2015年5月10日采用种子播种, 种植于甘肃农业大学与民乐县六坝镇诚泰药业有限公司板蓝根科研基地高新农业示范园, 试验地地处100° 43'28″ E, 38° 38'36″ N, 海拔 1824 m, 种子播种量30 kg· hm-2, 播种时在整好的畦面上按行距15 cm开挖3 cm的浅沟, 将种子均匀地播入沟内, 覆细土与畦面齐平, 保持土壤湿润, 播种后第6 天陆续出苗, 出苗后间苗, 留苗株距 7 cm左右, 田间管理同大田, 2015年不采收大青叶和板蓝根药材, 留株自然越冬繁殖种子。2016年越冬返青后从现蕾期开始田间观察测定, 并采用田间去雄、套袋及株间杂交等花朵处理及观察统计, 研究其开花习性和传粉方式。
从2016年4月20日菘蓝花蕾显现开始, 田间随机选取大小、长势相对一致, 无病虫害植株10株挂牌标记, 田间记录花序及小花在花序轴上的开放次序及持续时间, 参照任明迅[13]的方法, 选取具有代表性的10 枚花蕾, 观察其发育动态。通过采集不同等级的花蕾在显微镜下观察, 根据花药开裂程度和花粉成熟度的观测结果确定花药尚未成熟的花蕾标准, 作为去雄和套袋处理的花蕾。
现蕾期在田间随机选取大小、长势相对一致的菘蓝植株, 根据上述花蕾标准在田间选择发育一致的花蕾挂牌标记, 参照郭凤霞等[16]的方法采用套袋和不套袋, 人工授粉和自然授粉等8个处理(表1)。选择花萼微开缝, 但花瓣未露出花萼的花蕾操作处理, 此时柱头已成熟, 但花药尚未成熟开裂。套纸袋后第7 天摘除纸袋, 套网袋不需要摘除网袋。每一花序选择1~2朵发育良好的花蕾进行去雄操作, 其余花蕾全部摘除, 每处理重复操作20~60枚花蕾。去雄采用75%酒精消毒灭杀, 防除杂花粉污染, 去雄时若发现花蕾内有虫或个别花药开裂, 则摘除弃用, 重新选择操作。选择尼龙网(网孔1 mm× 1 mm)材料制作网袋(大小6 cm× 4 cm), 以严防昆虫传粉。选择防水透光的硫酸纸制作纸袋(大小6 cm× 4 cm)严防风媒传粉。去雄时保留部分花瓣, 待花瓣微露出花萼后授粉, 此时柱头活力较强, 并可避免蕾期自交的可能, 授粉后重新套袋。田间套袋时将袋从花序顶端套入, 用回形针夹住, 使袋隆起有一定空间, 以免束缚花朵生长发育, 造成花朵脱落。种子成熟时, 收集统计各处理短角果数和种子数, 最后计算结籽率和结实率, 并根据自交结种子数和结籽短角果数计算自交亲和指数(self-compatibility index, SCI)和自交不亲和指数(self-incompatibility index, SII)平均值[22]。采用王晓佳等[22]的自交亲和指数或自交不亲和指数标准判断菘蓝自交不亲和性, 即, 以0.5< SII≤ 1或0≤ SCI< 0.5为强自交不亲和性。采用郭凤霞等[16]和任跃英[27]的异交率(outcrossing rate, OCR)判断传粉方式, OCR< 5%为自花授粉, OCR> 50%为异花授粉, 5%< OCR< 50% 为常异花授粉。
结籽率(seed set rate)=结种子数/总花数× 100%
结实率(fruit rate)=结籽短角果总数/短角果总数× 100%
SCI=套纸袋自交结籽数/套纸袋总花蕾数(去雄同株异花人工授粉结籽数/去雄同株异花人工授粉总花蕾数)
SII=1-不去雄套纸袋自交结籽数/不去雄套纸袋总花蕾数(1-套纸袋自交结籽数/套纸袋自交总花蕾数)
| 表1 菘蓝花器不同处理及其检测目的 Table 1 Different treatments of floral organ and their detecting aims for I. indigotica |
从花蕾显现至开花结束, 在菘蓝试验田对传粉媒介观察, 从昆虫访花活动情况和花器操作传粉结实统计结果综合判定其传粉媒介。
采用Excel 2007统计软件绘图, 图中数据点为平均值± 标准误(mean± SE)。对于不同授粉处理的种子数、结籽率等性状指标的差异性采用单因素方差分析, 多重比较选用Duncan法。
田间观察发现, 民乐县栽培二倍体菘蓝属二年生草本药用植物。2015年采用种子播种, 2016年越冬返青后4月20日植株顶部陆续抽薹现蕾, 28日后花蕾陆续开放, 圆锥复总状花序, 生于枝顶(图1A, B)。开花后, 单株花序起初紧蹙在一起, 花序轴在开花的同时逐渐伸长, 花序自上而下依次开放, 首先主茎顶端的花序轴伸长, 顶端花序轴基部小花最先开放, 靠近顶枝的下一级花序轴小花随后开放, 单花序轴从花序的基部自下而上依次开放, 主茎顶部不断生出花序, 花序顶部不断生出小花蕾, 最后整个植株变得大而疏松, 形如锥形, 属于无限开花习性(图2A, B)。初现花蕾绿色, 发育过程中鲜黄色花瓣逐渐从花萼间隙显现伸长并展开。花朵开放与花药开裂均与气候环境条件有关, 当遇气温较低或阴雨天气时, 其开花延迟, 甚至不开放。无风晴朗天气条件下, 开花花朵花瓣向外张开角度大, 白天开花主要集中在上午8:00-12:00。遇暴风天气花瓣缩成半闭合状态。长雄蕊花药一般在开花20 min后花药开始开裂, 短雄蕊延后1~2 h才开裂, 花粉成熟时, 柱头与花药平齐, 随后柱头高度略低于长雄蕊花药, 略高于短雄蕊花药, 随着花的开放, 花丝拉伸至最长。开花盛期, 单株每天集中同时开放15~40朵, 每花序开放3~8朵。单株开花数、花蕾比和花朵开放率在单株间变异幅度较大, 变异系数均达100%以上, 花蕾比低于30%(表2)。
 | 图1 菘蓝开花期植株形态及花被特征 A: 大田群体现蕾期Field population bud emergence period; B: 大田群体开花期Field population anthesis.Fig.1 The flower shape and perianth characteristic of I. indigotica during blooming |
 | 图2 菘蓝花序特征及开花习性 A: 花序开花习性Flowering habit of inflorescences; B: 不同发育阶段的花序Different developing stages of inflorescences; C: 单花蕾Single flower bud; D: 单花正面Single flower front; E: 单花背面Single flower back; F: 单花侧面Single flower side; G: 单枚雌蕊Single pistil; H: 单枚雄蕊Single stamens.Fig.2 The inflorescence characteristic and flowering habit of I. indigotica |
| 表2 2016年4月28日民乐县菘蓝开花期田间单株开花统计 Table 2 Blooming statistics of single plant on April 28, 2016 in I. indigotica field in Minle county |
菘蓝主茎顶部着生花序和第一花序轴着生花数均最多, 沿主茎向下依次减少, 顶端花序轴着生花数60~150朵, 倒一级花序轴着生花数45~140朵, 倒二级花序轴着生花数20~100朵。初开花瓣呈十字180° 左右展开, 雄蕊围绕柱头60° 左右展开(图2C~F), 单花寿命6~7 d, 异常气候可缩短至3~4 d, 群体开花期30 d左右。雄蕊6枚, 4长2短, 淡黄色, 背着药, 纵裂且面向雌蕊开裂, 开放花朵的长、短雄蕊花丝长度相差约 1.00 mm左右; 子房上位近圆柱形, 柱头平截, 绿色, 雌蕊长2.30~2.62 mm(图2G, H)。
表3显示, 不同花器操作处理对菘蓝结籽率和结实率均具有极显著影响(P< 0.01)。在自然条件下, 菘蓝授粉结籽率和结实率均最高, 分别为95.00%和100.00%, 不去雄套纸袋不结种子, 不去雄套网袋结籽率较套纸袋提高5.0%, 结实率提高6.7%, 说明风媒也有微弱传粉作用。去雄套网袋较不去雄套网袋结籽率提高8.7%, 结实率提高7.6%, 进一步说明菘蓝存在风媒传粉概率。去雄套纸袋不结短角果和种子, 说明菘蓝不存在无融合生殖或孤雌生殖现象(parthenogenesis)。不去雄自然条件下, 菘蓝开花前套纸袋不结种子, 结籽率和结实率均为0, 揭示菘蓝存在较强的自交不亲和性。去雄套纸袋同株异花授粉结籽率和结实率均约4.0%, 去雄套纸袋异株异花授粉结籽率较自然授粉显著降低(P< 0.05), 结实率两者间的差异性不显著(P> 0.05), 但较同株异花结籽率提高68.0%(P< 0.01), 结实率提高93.0%以上(P< 0.01), 而去雄不套袋自然传粉条件下结籽率和结实率均在53.3%以上(表3), 说明菘蓝自花传粉力极弱, 同株异花结籽率很低, 显示出高度的自交不亲和性, 株间异交亲和性更强, 属异花授粉, 需传粉者, 昆虫为主要传粉媒介, 风媒也可辅助传粉。
| 表3 花器操作对菘蓝田间结实特性的影响 Table 3 Effect of floral organ operations on seed set traits for I. indigotica |
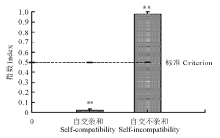
图3显示, 菘蓝自交不亲和指数变幅为0.96~1.00, 亲和指数变幅为0.00~0.04, 自然自交亲和指数为0, 人工去雄自交亲和指数0.04, 平均自交亲和指数(SCI)极显著小于0.5(P< 0.01)。菘蓝自然自交不亲和指数为1, 人工自交不亲和指数0.96, 平均自交不亲和指数(SII)接近1, 极显著大于0.5(P< 0.01), 按照王晓佳等[22]的标准判断, 属于典型的自交不亲和性植物。
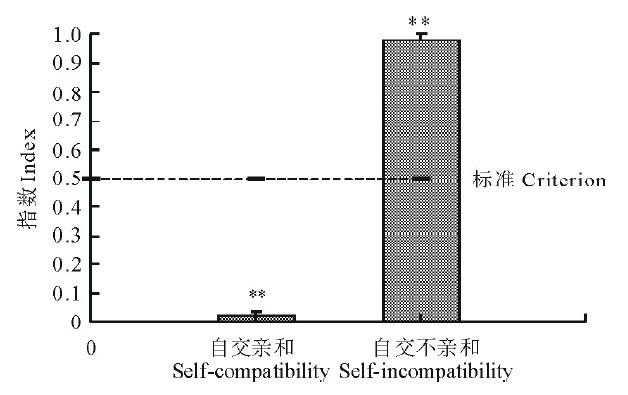 | 图3 菘蓝自交亲和指数与自交不亲和指数比较 * * 表示与标准(0.5)相比存在极显著差异(P< 0.01)。* * indicate there has significant difference compore with criterion (0.5) (P< 0.01).Fig.3 Comparison of self-compatibility index and self-incompatibility index for I. indigotica |
田间观察发现, 菘蓝在花蕾期访花昆虫数量多, 活动频繁, 在花序间穿梭, 这些昆虫在菘蓝植株上往返访花采食花粉(图4), 在植株花序上访花停留时间随植株和花序的不同而异。初步鉴定访花昆虫主要是食蚜蝇(Scaeva pyrastri)(图4A, B)、黑蜂(Apis mellifer)(图4C)。这些昆虫的胸、腹、足等部位置显微镜下抖落观察均粘有大量的花粉粒。田间试验条件下, 当微风吹过时, 开放花朵的花冠中散落大量花粉粒, 但遇大风天气时, 花瓣几乎处于闭合状态, 不利于风力向外散布花粉, 说明昆虫是菘蓝实现有性繁殖的主要传粉媒介, 风媒只起到辅助传粉的媒介作用。
 | 图4 菘蓝花朵间传粉昆虫活动情况 A-B: 食蚜蝇Episyrphus balteatus; C: 黑蜂Apis melliferFig.4 Activity conditions of pollination insects among I. indigotica flowers |
植物繁育系统是指能够代表直接影响其后代遗传组成的所有有性特征[23], 主要包括植物花部特征、花器官寿命、花开放式样、自交亲和程度和交配系统[24]。野生药用植物高乌头(Aconitum sinomontanum)主茎自下而上依次递进开花[25]。开花习性也是物种长期适应环境的一种保护策略。本研究发现, 二倍体菘蓝生长第2年植株较大、群体开花期持续30 d, 花小而密集, 单株内花序与同一花序轴上花朵开放次序异向, 花序从株顶自上而下, 而单花序内自下而上依次开放, 形成明显反向开放次序, 这种无限开花习性有利于吸引昆虫, 同时也可降低鸟类啄食, 顶部花蕾可阻挡强光和大风, 在一定程度上保护开放花朵。初现绿色花蕾花药尚未成熟开裂, 说明闭花授粉受限。花瓣微露出花萼后开花当天, 花粉量最多, 雄蕊的长短不齐也反映出占据空间位置的效能。花瓣, 雄蕊和花丝均呈鲜黄色, 由于黄色能吸收紫外光, 可避免花器受到强紫外伤害, 黄色也可扮演花粉和花粉囊色, 吸引昆虫, 提高传粉效率, 显花植物为了吸引传粉昆虫而排斥食花害虫访花的决定, 利用花特有的颜色和气味行使信号功能[16], 贝母属植物黄花(Fritillaria koidzuminia)可吸引蜜蜂和蝇类等传粉者[26], 这也反映出菘蓝虫媒传粉的实事。田间观察也发现, 菘蓝盛花期, 有许多蜂类、蝇类等昆虫访花, 白天随气温升高数量增多, 从上午10:00至12:00和下午2:00至3:00, 访花者数量出现峰值, 田间访花活动最为频繁的昆虫主要是食蚜蝇和黑蜂, 访花者在某一株上采食花粉后, 身体绒毛粘有大量花粉, 又飞往另一株采食, 大多数采食花朵花瓣微开, 花粉鲜黄, 进一步印证了昆虫传粉的现实。但是并不能排除风媒传粉的可能, 因为鲜黄的花粉多而散, 随风散落花器四周, 这一结果进一步证明菘蓝传粉媒介以虫媒为主, 风媒为辅。
植物传粉受精过程与种子的形成和质量具有重要的关系, 是高等植物传宗接代过程中最主要的环节, 在植物繁殖、遗传、变异、进化等方面具有重要的作用[23, 24]。在自然界中植物开花后成熟花粉在传粉媒介作用下从雄蕊花药或小包子囊中散出后传送到雌蕊柱头或胚珠上的过程称为传粉, 传粉媒介主要有蜜蜂、甲虫、蝇类等和风, 此外蝙蝠、蜗牛也能传粉, 有些植物还通过水进行传粉[23]。药用植物育种中, 根据花部结构、开花习性和传粉特性等方面分为3种类型:异交率< 5%为自花授粉, 异交率> 50%为异花授粉, 异交率介于 5%~50% 为常异花授粉[16, 27]。测定植物亲和指数一般在花期采取密网阻隔飞行昆虫、用化学药剂阻隔爬行类昆虫、套袋隔离和增大株距等措施, 结籽还受植株营养状况、花粉活力、柱头可授性、花枝部位和气候(光照、温度和水分)等生育差异带来的影响[27]。菘蓝是十字花科药用植物, 解决菘蓝制种的技术问题是杂种优势利用的关键。本研究2016年在民乐县大田采用两年生的菘蓝进行套袋隔离花器操作处理方法, 为了减少误差, 通过在授粉前对花粉活力和柱头可授性预检测, 选择花枝部位分枝和花期一致的花蕾, 研究结果发现在4月中旬菘蓝现蕾开始柱头已具有可授性, 但花药未开裂, 不能闭花授粉, 此期去雄套纸袋处理不结实, 说明本研究选蕾恰当, 也说明菘蓝不存在无融合生殖现象, 这与胡春等[28]对青藏高原优势毒杂草钝裂银莲花(Anemone obtusiloba)研究的结论相反, 反映出毒草自繁能力更强, 因为郭凤霞等[16]研究也发现, 濒危珍稀药用植物暗紫贝母不存在无融合生殖现象。本研究还揭示, 菘蓝自然授粉结籽率为95.0%, 去雄套纸袋异株异花人工授粉结籽率72.0%, 去雄不套袋结籽率53.3%, 不去雄套网袋结籽率5.0%, 去雄后套网袋提高至13.7%, 说明菘蓝属于典型异花授粉类型, 去雄套纸袋异花人工授粉低于自然授粉, 可能是由于菘蓝花朵小, 纸袋隔绝风媒传粉概率, 也可能是套纸袋对雌蕊还有遮阴扰动对去雄后花器受损的补偿效应在一定程度上损耗了雌蕊营养。套网袋条件下, 去雄结籽率较不去雄后提高8.7%, 说明雄蕊还具有一定的传粉物理隔离作用。菘蓝不去雄和去雄套纸袋均不结实, 去雄套纸袋人工同株异花授粉结籽率和结实率仅4%左右, 既反映出菘蓝风媒传粉的可能, 也反映出菘蓝较强的自交不亲和性, 这与高延辉等[6, 17]对吉林长春市二年生菘蓝显微镜检得出的结论相一致, 即菘蓝在花期套袋天然授粉的自交亲和指数为0.078, 花期人工授粉的自交亲和指数为0.110, 不同的是本研究在民乐县自然气候条件下套袋天然授粉的自交亲和指数为0, 花期人工授粉的自交亲和指数仅0.040, 揭示菘蓝在该区自然气候条件下自交不亲和性表现更强, 这种特性可能是适宜环境条件下菘蓝回避自交退化的一种本能的应激反应, 因为高寒区濒危药用植物暗紫贝母具有弱的自交亲和性, 通过部分自交亲和提高极端环境条件下花粉传递效率, 才使得稀有物种得以繁衍生息[16], 但民乐县属于河西走廊地区, 光、温和水资源丰富, 长期温湿度适宜的生态压力下不利于菘蓝自花传粉行为的完成, 而有助于株间花粉竞争传粉行为的驱动。
菘蓝株内具有异向无限开花习性, 不存在无融合生殖, 具有较强的自交不亲和性, 异花传粉效率高, 访花者主要有蜂类、蝇类等, 具有培育优良自交不亲和系进行杂种优势利用的巨大潜力, 甘肃省河西走廊地区民乐县一带是菘蓝制种的优势生态区, 对菘蓝传粉蜜蜂种类及访花频率的研究、菘蓝自交不亲和系的选育和杂交种的配置将是下一步研究的重点, 菘蓝开花期长, 可在开发菘蓝花蜂蜜产业中引起关注, 以期促进菘蓝的产业化健康发展。
致谢:甘肃省民乐县诚泰药业有限公司韩大雷提供试验地和帮助, 成都天地网信息科技有限公司的张斌参与田间管理。本科生张荩凤、徐博琼、王柏林、王雪琴、张旭鹏、周艳参与试验指标测定, 在此一并致谢。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|